

The visual pigment xenopsin is widespread in protostome eyes and impacts the view on eye evolution
- PMID: 32880369
- PMCID: PMC7529461
- DOI: 10.7554/eLife.55193
The visual pigment xenopsin is widespread in protostome eyes and impacts the view on eye evolution
Abstract
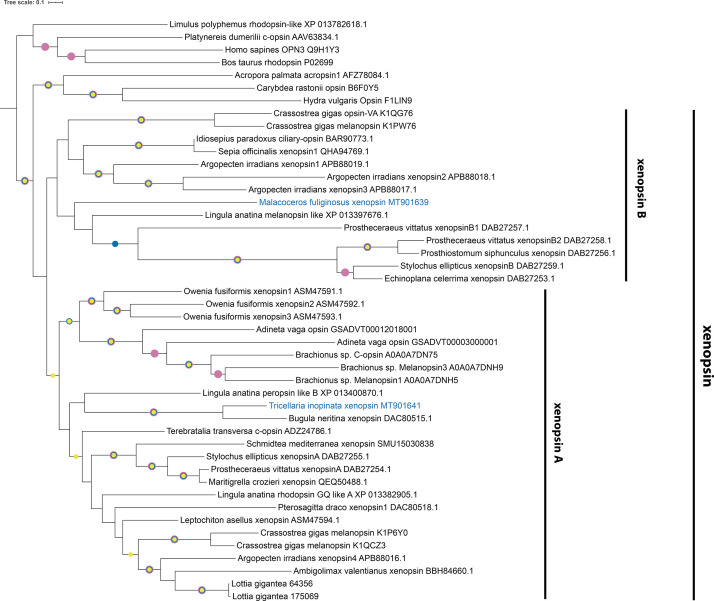
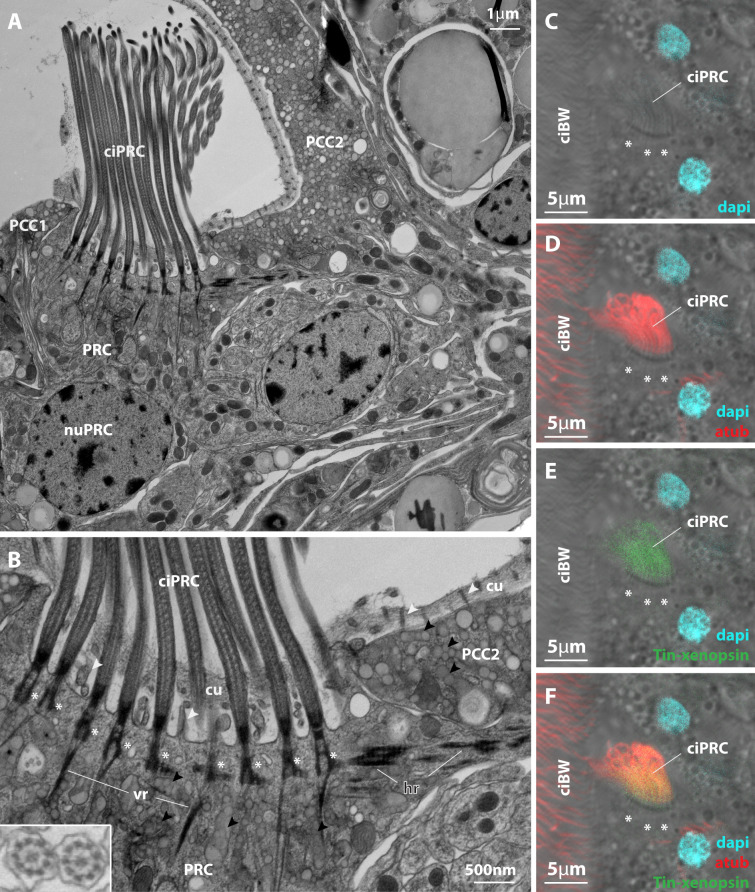
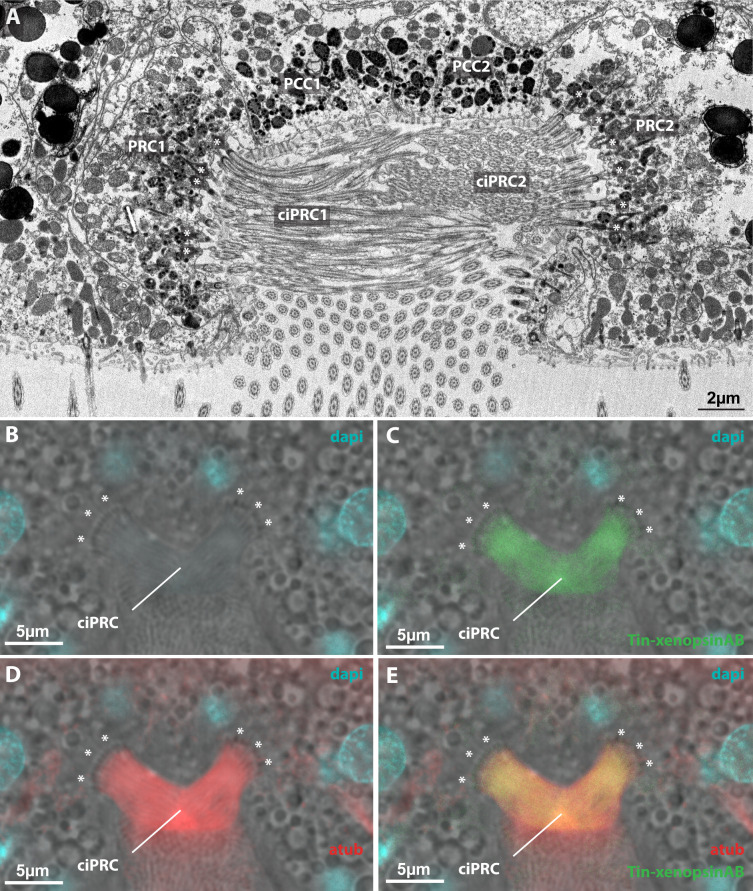
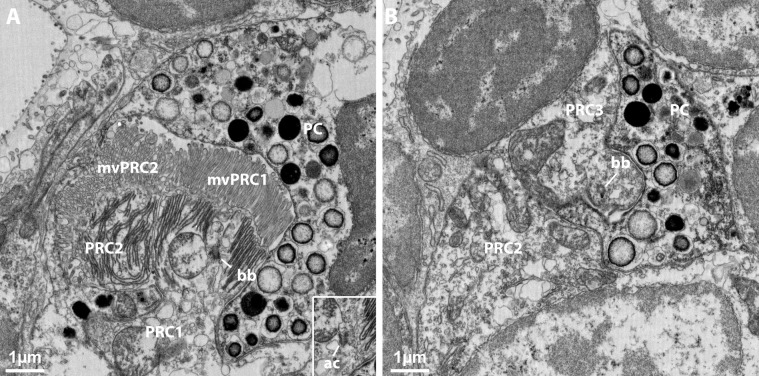
Photoreceptor cells in the eyes of Bilateria are often classified into microvillar cells with rhabdomeric opsin and ciliary cells with ciliary opsin, each type having specialized molecular components and physiology. First data on the recently discovered xenopsin point towards a more complex situation in protostomes. In this study, we provide clear evidence that xenopsin enters cilia in the eye of the larval bryozoan Tricellaria inopinata and triggers phototaxis. As reported from a mollusc, we find xenopsin coexpressed with rhabdomeric-opsin in eye photoreceptor cells bearing both microvilli and cilia in larva of the annelid Malacoceros fuliginosus. This is the first organism known to have both xenopsin and ciliary opsin, showing that these opsins are not necessarily mutually exclusive. Compiling existing data, we propose that xenopsin may play an important role in many protostome eyes and provides new insights into the function, evolution, and possible plasticity of animal eye photoreceptor cells.
Keywords: evolution; evolutionary biology; eyes; malacoceros fuliginosus; neuroscience; opsins; photoreceptors; tricellaria inopinata.
© 2020, Döring et al.
Conflict of interest statement
CD, SK, ST, IK, HH No competing interests declared
Figures


















Similar articles
-
Co-expression of xenopsin and rhabdomeric opsin in photoreceptors bearing microvilli and cilia.Elife. 2017 Sep 6;6:e23435. doi: 10.7554/eLife.23435. Elife. 2017. PMID: 28876222 Free PMC article.
-
Extraocular, rod-like photoreceptors in a flatworm express xenopsin photopigment.Elife. 2019 Oct 22;8:e45465. doi: 10.7554/eLife.45465. Elife. 2019. PMID: 31635694 Free PMC article.
-
Expression dynamics and protein localization of rhabdomeric opsins in Platynereis larvae.Integr Comp Biol. 2013 Jul;53(1):7-16. doi: 10.1093/icb/ict046. Epub 2013 May 10. Integr Comp Biol. 2013. PMID: 23667045 Free PMC article.
-
Evolution of eyes and photoreceptor cell types.Int J Dev Biol. 2003;47(7-8):563-71. Int J Dev Biol. 2003. PMID: 14756332 Review.
-
Reconstructing the eyes of Urbilateria.Philos Trans R Soc Lond B Biol Sci. 2001 Oct 29;356(1414):1545-63. doi: 10.1098/rstb.2001.0971. Philos Trans R Soc Lond B Biol Sci. 2001. PMID: 11604122 Free PMC article. Review.
Cited by
-
Deep Diversity: Extensive Variation in the Components of Complex Visual Systems across Animals.Cells. 2022 Dec 8;11(24):3966. doi: 10.3390/cells11243966. Cells. 2022. PMID: 36552730 Free PMC article. Review.
-
A chromosome-level genome assembly of the disco clam, Ctenoides ales.G3 (Bethesda). 2024 Sep 4;14(9):jkae115. doi: 10.1093/g3journal/jkae115. G3 (Bethesda). 2024. PMID: 38805695 Free PMC article.
-
Molluscan Genomes Reveal Extensive Differences in Photopigment Evolution Across the Phylum.Mol Biol Evol. 2023 Dec 1;40(12):msad263. doi: 10.1093/molbev/msad263. Mol Biol Evol. 2023. PMID: 38039155 Free PMC article.
-
Convergent evolutionary counterion displacement of bilaterian opsins in ciliary cells.Cell Mol Life Sci. 2022 Aug 24;79(9):493. doi: 10.1007/s00018-022-04525-6. Cell Mol Life Sci. 2022. PMID: 36001156 Free PMC article.
-
Duplication and Losses of Opsin Genes in Lophotrochozoan Evolution.Mol Biol Evol. 2023 Apr 4;40(4):msad066. doi: 10.1093/molbev/msad066. Mol Biol Evol. 2023. PMID: 36947081 Free PMC article.
References
-
- Applebury ML, Antoch MP, Baxter LC, Chun LL, Falk JD, Farhangfar F, Kage K, Krzystolik MG, Lyass LA, Robbins JT. The murine cone photoreceptor: a single cone type expresses both S and M opsins with retinal spatial patterning. Neuron. 2000;27:513–523. doi: 10.1016/s0896-6273(00)00062-3. - DOI - PubMed
-
- Arendt D, Tessmar K, de Campos-Baptista MI, Dorresteijn A, Wittbrodt J. Development of pigment-cup eyes in the polychaete Platynereis dumerilii and evolutionary conservation of larval eyes in bilateria. Development. 2002;129:1143–1154. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
