

A connectome and analysis of the adult Drosophila central brain
- PMID: 32880371
- PMCID: PMC7546738
- DOI: 10.7554/eLife.57443
A connectome and analysis of the adult Drosophila central brain
Abstract
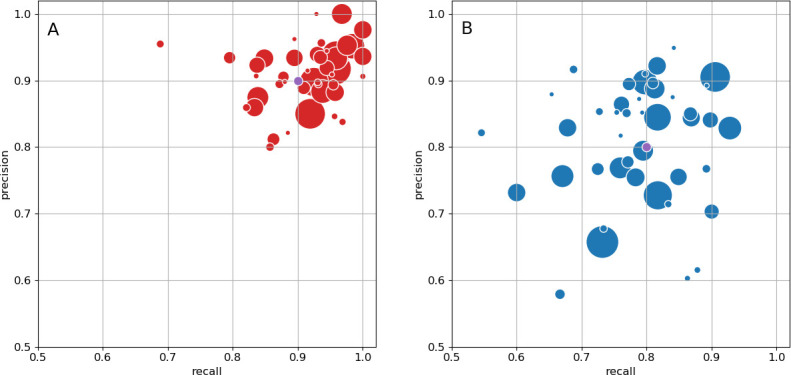
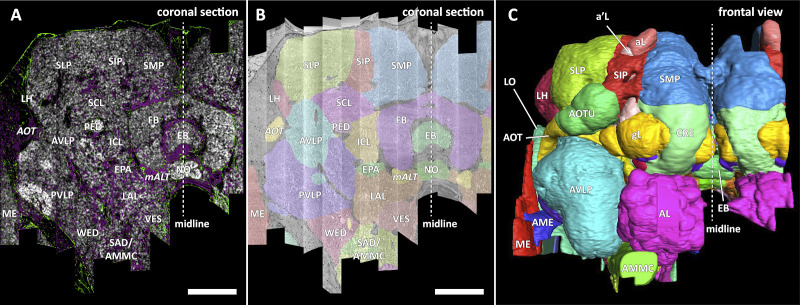
The neural circuits responsible for animal behavior remain largely unknown. We summarize new methods and present the circuitry of a large fraction of the brain of the fruit fly Drosophila melanogaster. Improved methods include new procedures to prepare, image, align, segment, find synapses in, and proofread such large data sets. We define cell types, refine computational compartments, and provide an exhaustive atlas of cell examples and types, many of them novel. We provide detailed circuits consisting of neurons and their chemical synapses for most of the central brain. We make the data public and simplify access, reducing the effort needed to answer circuit questions, and provide procedures linking the neurons defined by our analysis with genetic reagents. Biologically, we examine distributions of connection strengths, neural motifs on different scales, electrical consequences of compartmentalization, and evidence that maximizing packing density is an important criterion in the evolution of the fly's brain.
Keywords: D. melanogaster; brain regions; cell types; computational biology; connectome; connectome reconstuction methods; graph properties; neuroscience; synapse detecton; systems biology.
Plain language summary
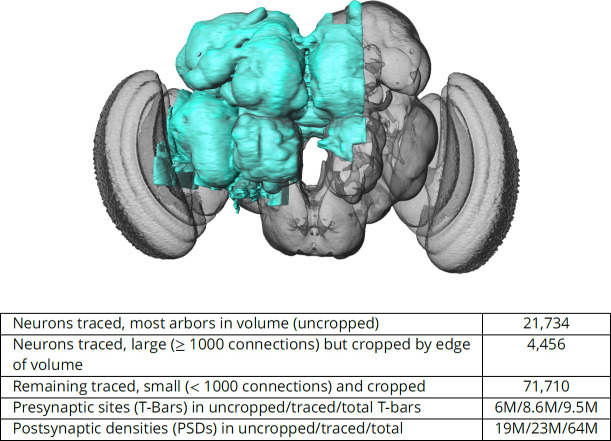
Animal brains of all sizes, from the smallest to the largest, work in broadly similar ways. Studying the brain of any one animal in depth can thus reveal the general principles behind the workings of all brains. The fruit fly Drosophila is a popular choice for such research. With about 100,000 neurons – compared to some 86 billion in humans – the fly brain is small enough to study at the level of individual cells. But it nevertheless supports a range of complex behaviors, including navigation, courtship and learning. Thanks to decades of research, scientists now have a good understanding of which parts of the fruit fly brain support particular behaviors. But exactly how they do this is often unclear. This is because previous studies showing the connections between cells only covered small areas of the brain. This is like trying to understand a novel when all you can see is a few isolated paragraphs. To solve this problem, Scheffer, Xu, Januszewski, Lu, Takemura, Hayworth, Huang, Shinomiya et al. prepared the first complete map of the entire central region of the fruit fly brain. The central brain consists of approximately 25,000 neurons and around 20 million connections. To prepare the map – or connectome – the brain was cut into very thin 8nm slices and photographed with an electron microscope. A three-dimensional map of the neurons and connections in the brain was then reconstructed from these images using machine learning algorithms. Finally, Scheffer et al. used the new connectome to obtain further insights into the circuits that support specific fruit fly behaviors. The central brain connectome is freely available online for anyone to access. When used in combination with existing methods, the map will make it easier to understand how the fly brain works, and how and why it can fail to work correctly. Many of these findings will likely apply to larger brains, including our own. In the long run, studying the fly connectome may therefore lead to a better understanding of the human brain and its disorders. Performing a similar analysis on the brain of a small mammal, by scaling up the methods here, will be a likely next step along this path.
© 2020, Scheffer et al.
Conflict of interest statement
LS, CX, ZL, ST, KH, GH, KS, SB, JC, PH, WK, LU, TZ, DA, JB, TD, DK, TK, KK, NN, DO, HO, ET, MI, AB, JG, TW, RS, PS, EN, CK, CA, DB, SB, JB, BC, NC, MC, MD, OD, BE, KF, SF, NF, AF, GH, EJ, SK, NK, JK, SL, AL, CM, EM, SM, CM, MN, OO, NO, CO, NP, CP, TP, EP, EP, NR, CR, MR, JR, SR, MS, AS, AS, AS, CS, KS, NS, MS, AS, JS, ST, IT, DT, ET, TT, JW, TY, JH, FL, RP, PR, VJ, MC, GJ, KI, SS, RG, IM, GR, HH, SP No competing interests declared, MJ, JM, TB, LL, PL, LL, VJ is an employee of Google.
Figures


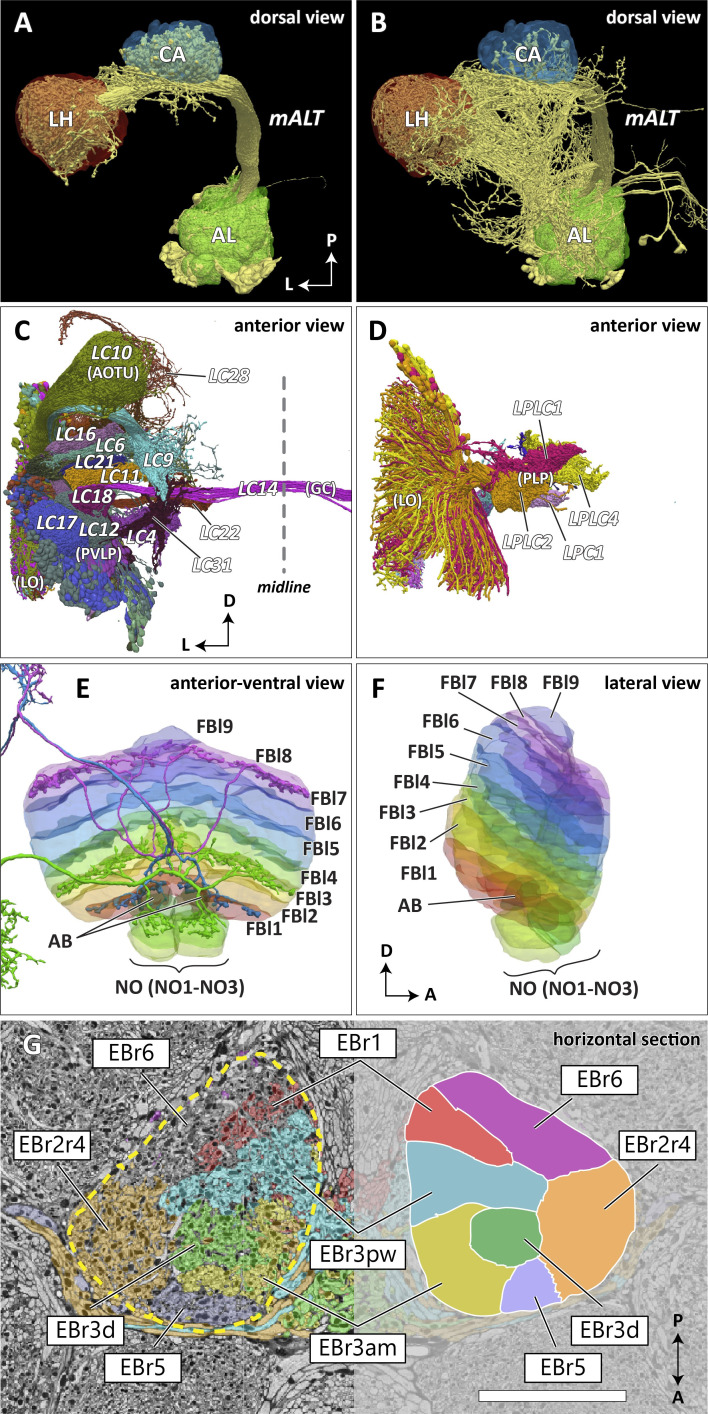
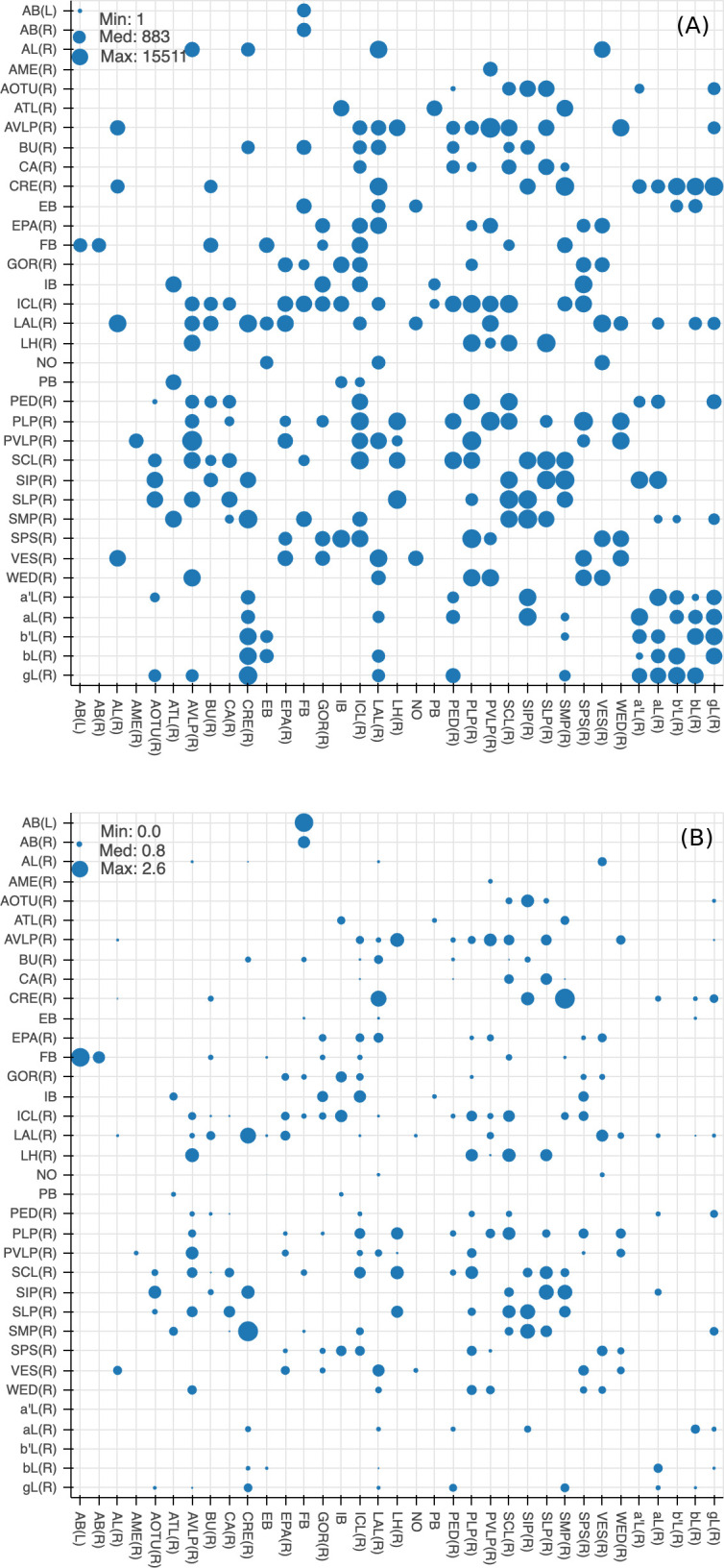
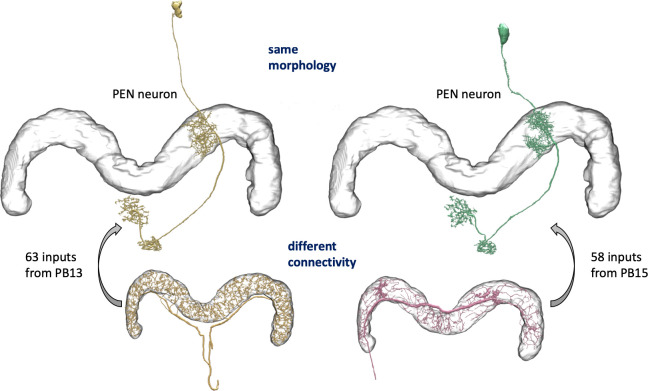
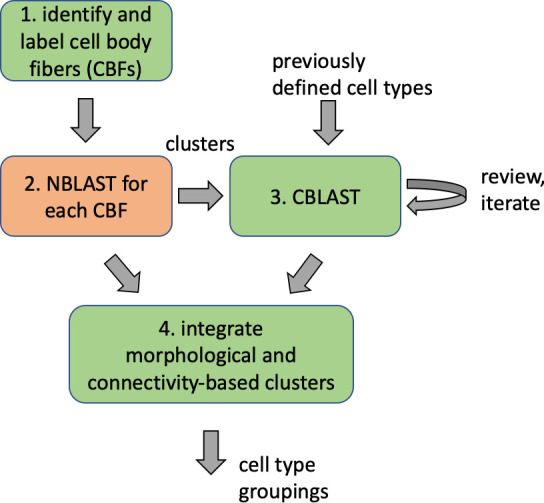
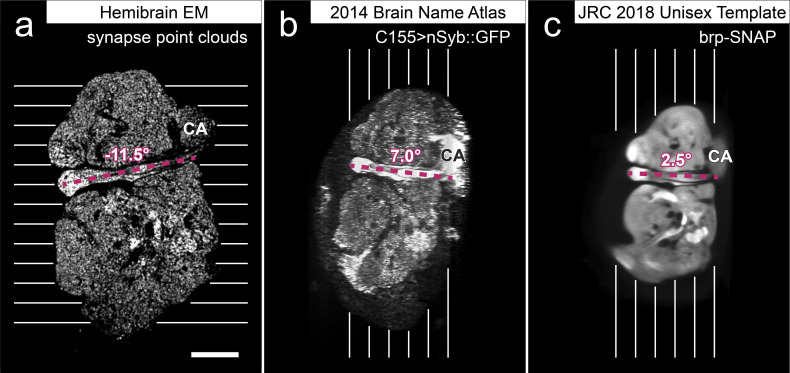
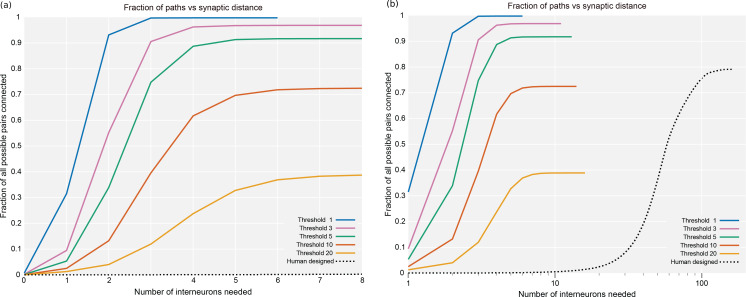
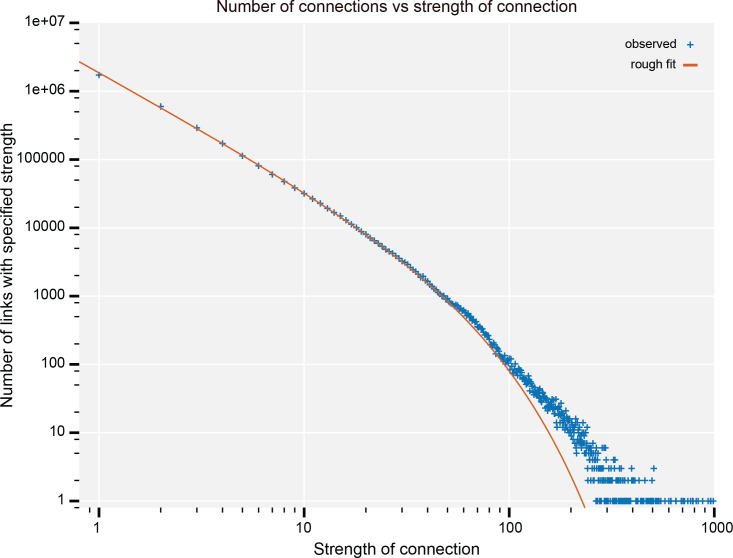
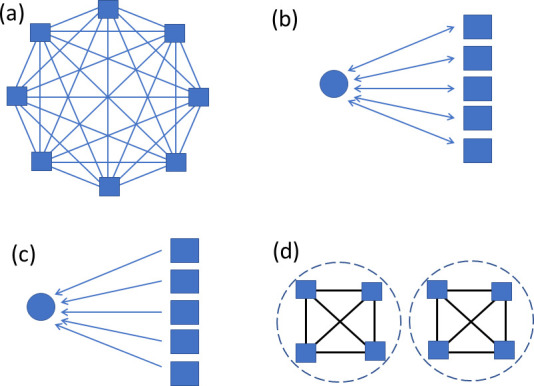
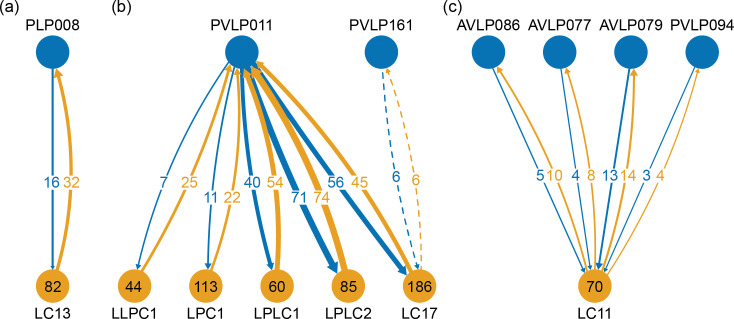
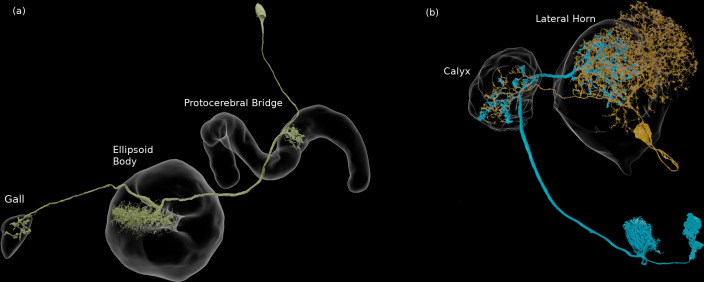
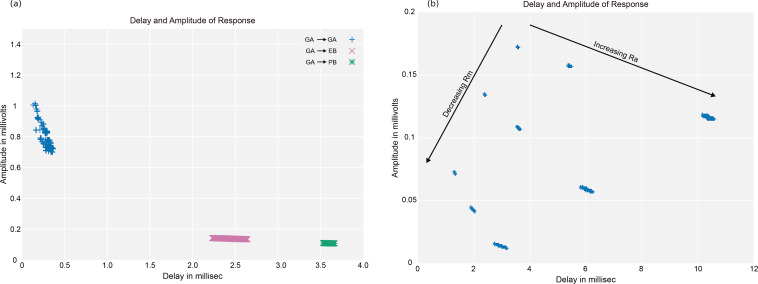
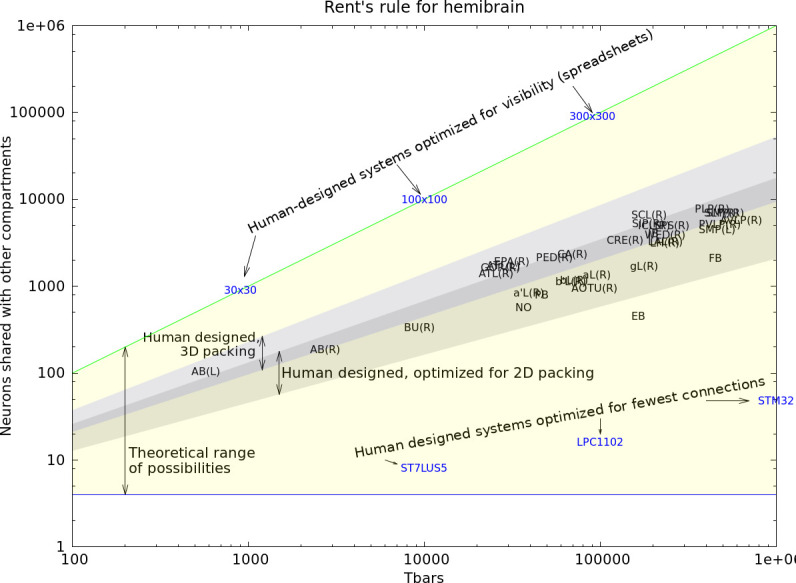
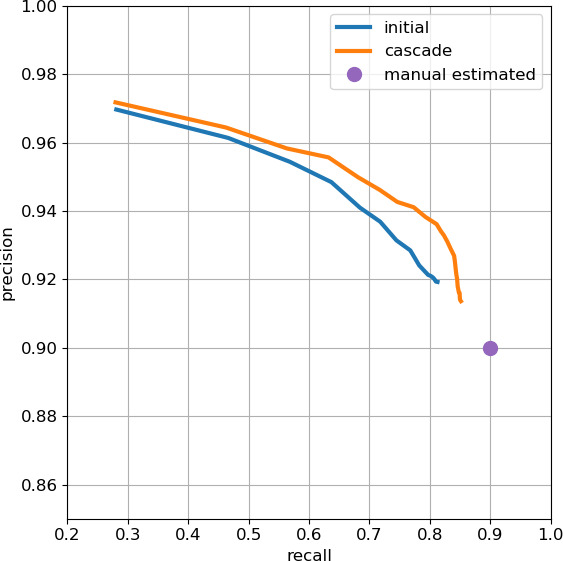
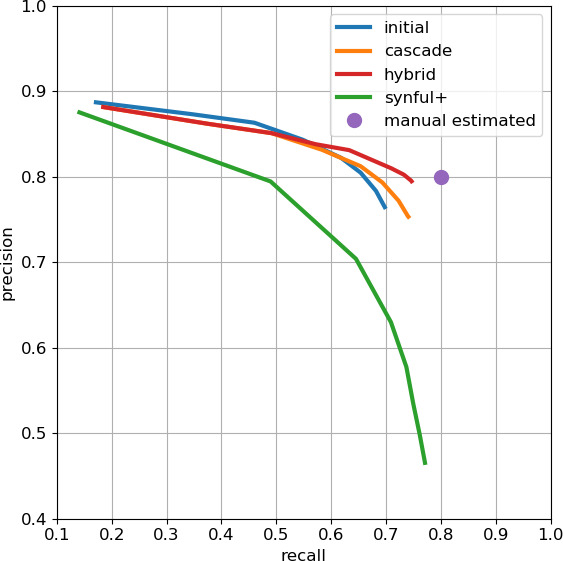
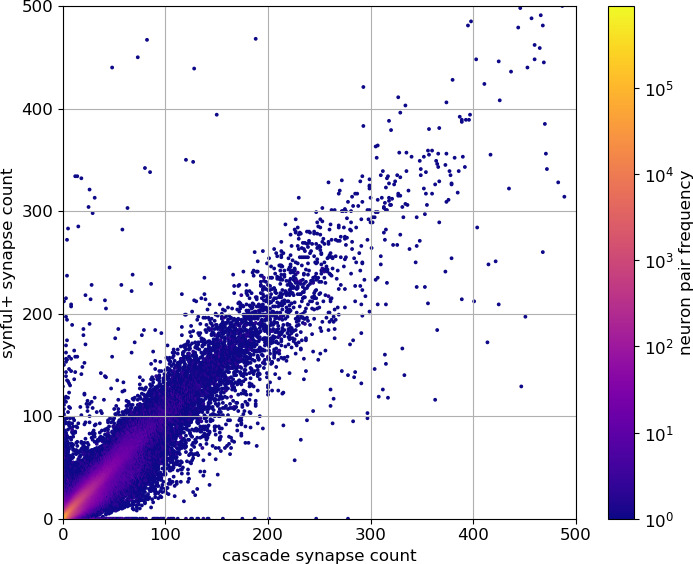
Comment in
-
Mapping the mind of a fly.Elife. 2020 Oct 8;9:e62451. doi: 10.7554/eLife.62451. Elife. 2020. PMID: 33030427 Free PMC article.
References
-
- Ascoli GA, Donohue DE, Halavi M. NeuroMorpho.Org: a central resource for neuronal morphologies. Journal of Neuroscience. 2007;27:9247–9251. doi: 10.1523/JNEUROSCI.2055-07.2007. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
