

Actomyosin Contractility in the Generation and Plasticity of Axons and Dendritic Spines
- PMID: 32882840
- PMCID: PMC7565476
- DOI: 10.3390/cells9092006
Actomyosin Contractility in the Generation and Plasticity of Axons and Dendritic Spines
Abstract
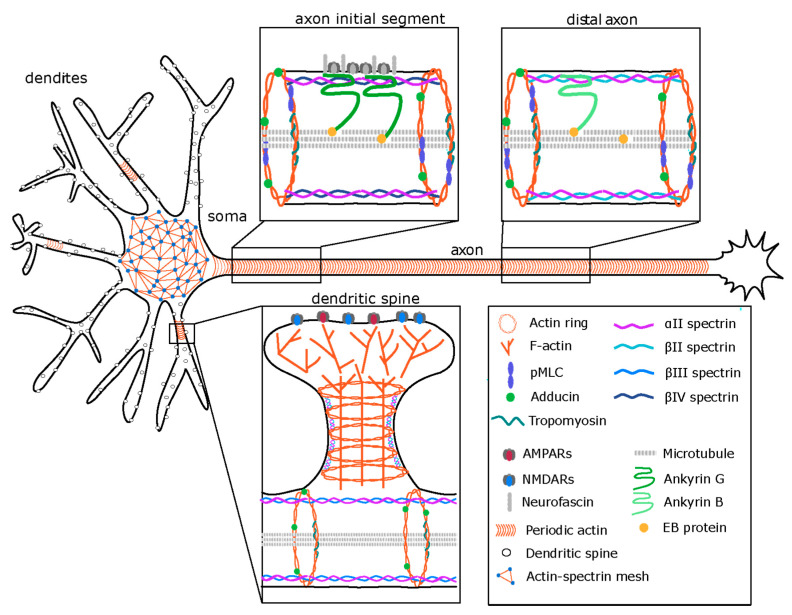
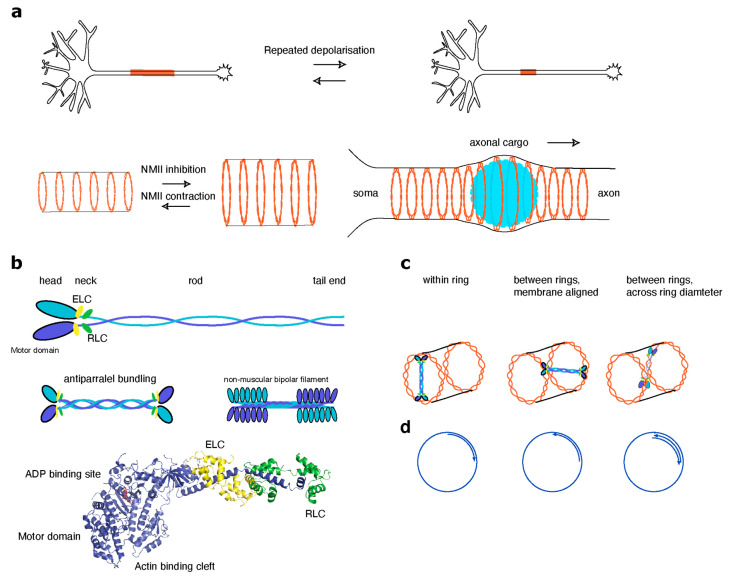
Actin and non-muscle myosins have long been known to play important roles in growth cone steering and neurite outgrowth. More recently, novel functions for non-muscle myosin have been described in axons and dendritic spines. Consequently, possible roles of actomyosin contraction in organizing and maintaining structural properties of dendritic spines, the size and location of axon initial segment and axonal diameter are emerging research topics. In this review, we aim to summarize recent findings involving myosin localization and function in these compartments and to discuss possible roles for actomyosin in their function and the signaling pathways that control them.
Keywords: AIS; actin; calcium signaling; calpain; dendritic spines; myosin; spectrin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Costa A.C.R., Sousa S.C., Pinto-Costa R., Mateus J.C., Lopes C.D.F., Costa A.C.R., Rosa D., Machado D., Pajuelo L., Wang X., et al. The membrane periodic skeleton is an actomyosin network that regulates axonal diameter and conduction. eLife. 2020;9:1–20. doi: 10.7554/eLife.55471. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
