

Fine-tuning sugar content in strawberry
- PMID: 32883370
- PMCID: PMC7470447
- DOI: 10.1186/s13059-020-02146-5
Fine-tuning sugar content in strawberry
Erratum in
-
Correction: Fine-tuning sugar content in strawberry.Genome Biol. 2025 Dec 24;26(1):435. doi: 10.1186/s13059-025-03894-y. Genome Biol. 2025. PMID: 41444675 Free PMC article. No abstract available.
Abstract
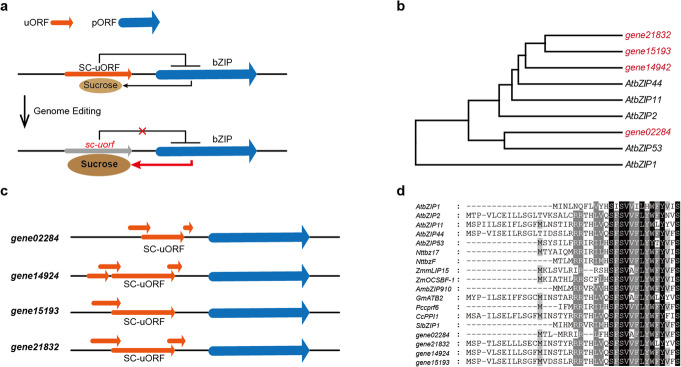
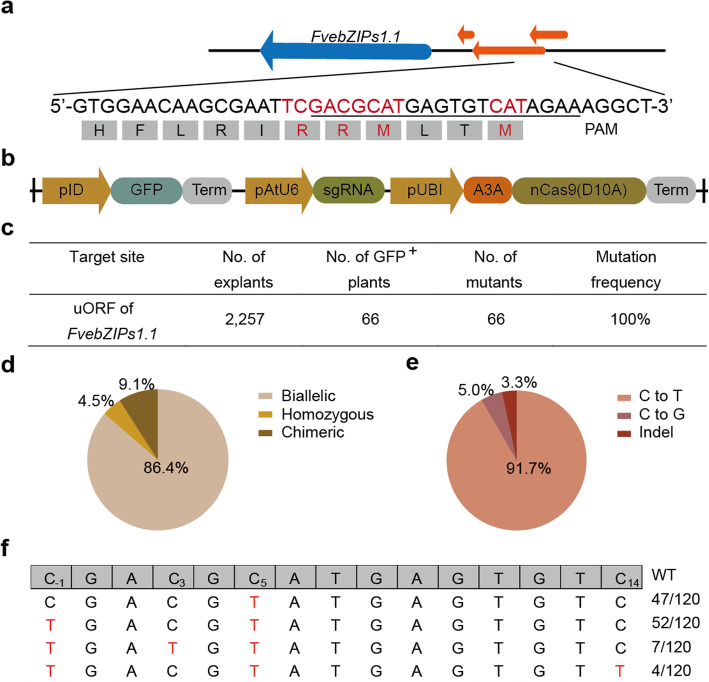
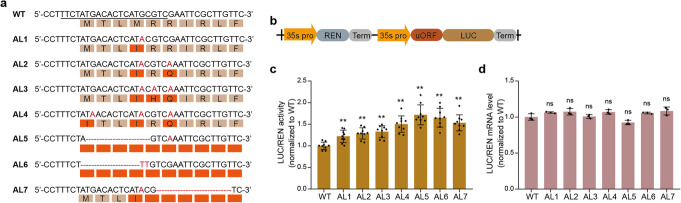
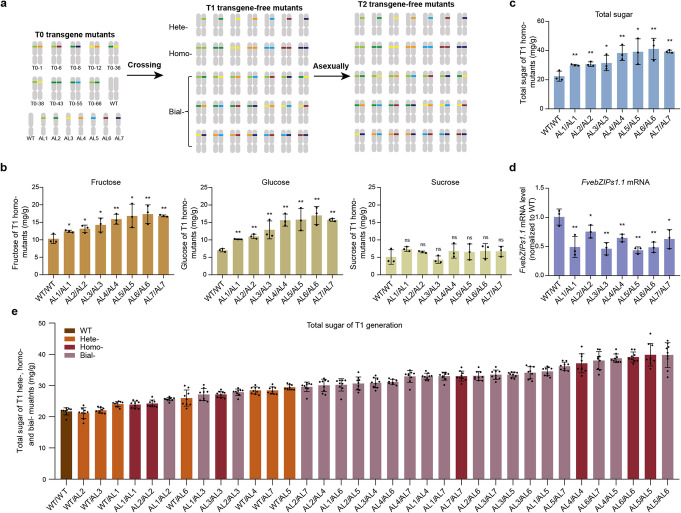
Fine-tuning quantitative traits for continuous subtle phenotypes is highly advantageous. We engineer the highly conserved upstream open reading frame (uORF) of FvebZIPs1.1 in strawberry (Fragaria vesca), using base editor A3A-PBE. Seven novel alleles are generated. Sugar content of the homozygous T1 mutant lines is 33.9-83.6% higher than that of the wild-type. We also recover a series of transgene-free mutants with 35 novel genotypes containing a continuum of sugar content. All the novel genotypes could be immediately fixed in subsequent generations by asexual reproduction. Genome editing coupled with asexual reproduction offers tremendous opportunities for quantitative trait improvement.
Keywords: Asexually reproducing crops; Basic leucine zipper; Fine-tuning; Quantitative trait variation; Strawberry; Sugar content; Upstream open reading frame.
Conflict of interest statement
None
Figures
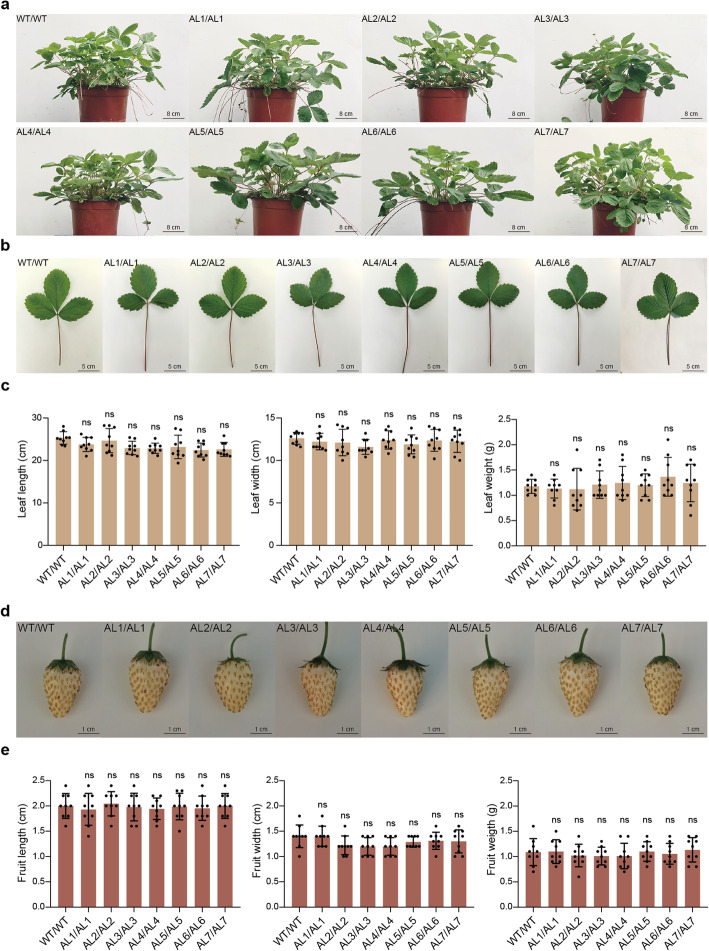
Comment in
-
Base edit your way to better crops.Nature. 2022 Apr;604(7907):790-792. doi: 10.1038/d41586-022-01117-z. Nature. 2022. PMID: 35478229 No abstract available.
References
-
- Rodriguez-Leal D, Lemmon ZH, Man J, Bartlett ME, Lippman ZB. Engineering quantitative trait variation for crop improvement by genome editing. Cell. 2017;171:470–480. - PubMed
-
- McKey D, Elias M, Pujol B, Duputie A. The evolutionary ecology of clonally propagated domesticated plants. New Phytol. 2010;186:318–332. - PubMed
-
- Schoof H, Lenhard M, Haecker A, Mayer KFX, Jurgens G, Laux T. The stem cell population of Arabidopsis shoot meristems is maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell. 2000;100:635–644. - PubMed
-
- Vrebalov J, Ruezinsky D, Padmanabhan V, White R, Medrano D, Drake R, Schuch W, Giovannoni J. A MADS-box gene necessary for fruit ripening at the tomato ripening-inhibitor (Rin) locus. Science. 2002;296:343–346. - PubMed
-
- Xu C, Liberatore KL, MacAlister CA, Huang ZJ, Chu YH, Jiang K, Brooks C, Ogawa-Ohnishi M, Xiong GY, Pauly M, et al. A cascade of arabinosyltransferases controls shoot meristem size in tomato. Nat Genet. 2015;47:784–792. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
