

Iron at the host-microbe interface
- PMID: 32883564
- PMCID: PMC7554189
- DOI: 10.1016/j.mam.2020.100895
Iron at the host-microbe interface
Abstract
Iron is an essential micronutrient for nearly all living organisms. In addition to facilitating redox reactions, iron is bound by metalloproteins that participate in a variety of biological processes. As the bioavailability of free iron in host environments is extremely low, iron lies at the center of a battle for nutrients between microbes and their host. Mucosal surfaces such as the respiratory and gastrointestinal tracts are constantly exposed to commensal and pathogenic microorganisms. Whereas a key strategy of mammalian antimicrobial defense is to deprive microbes of iron, pathogens and some commensals have evolved effective strategies to circumvent iron limitation. Here we provide an overview of mechanisms underpinning the tug-of-war for iron between microbes and their host, with a particular focus on mucosal surfaces.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Figures
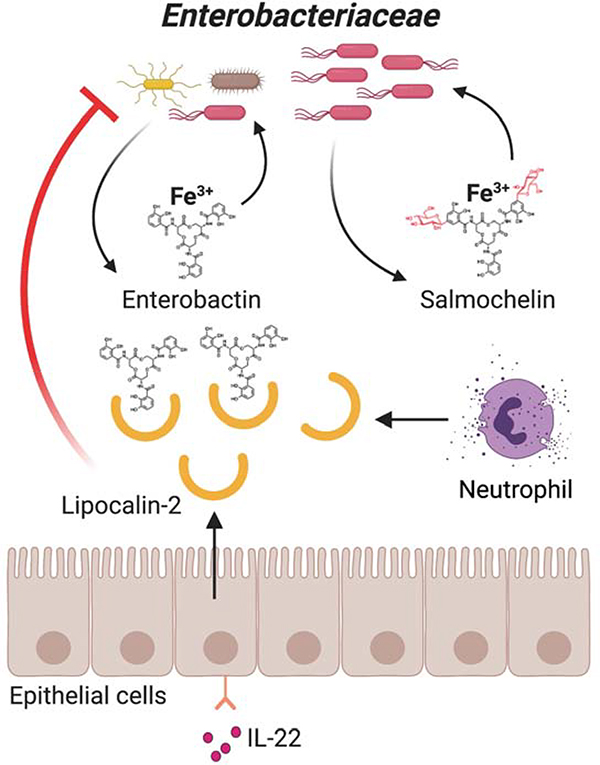
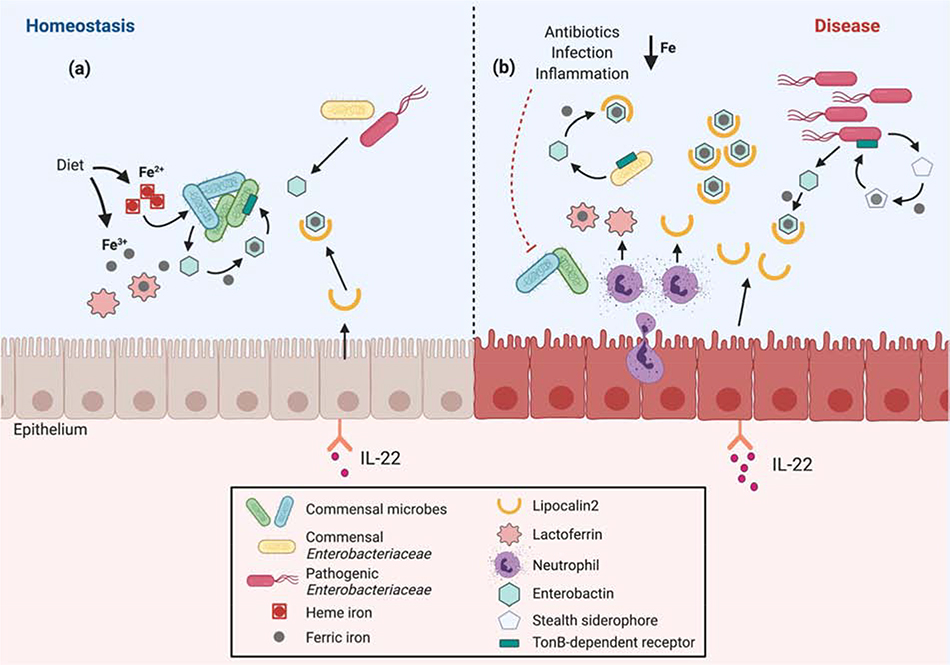
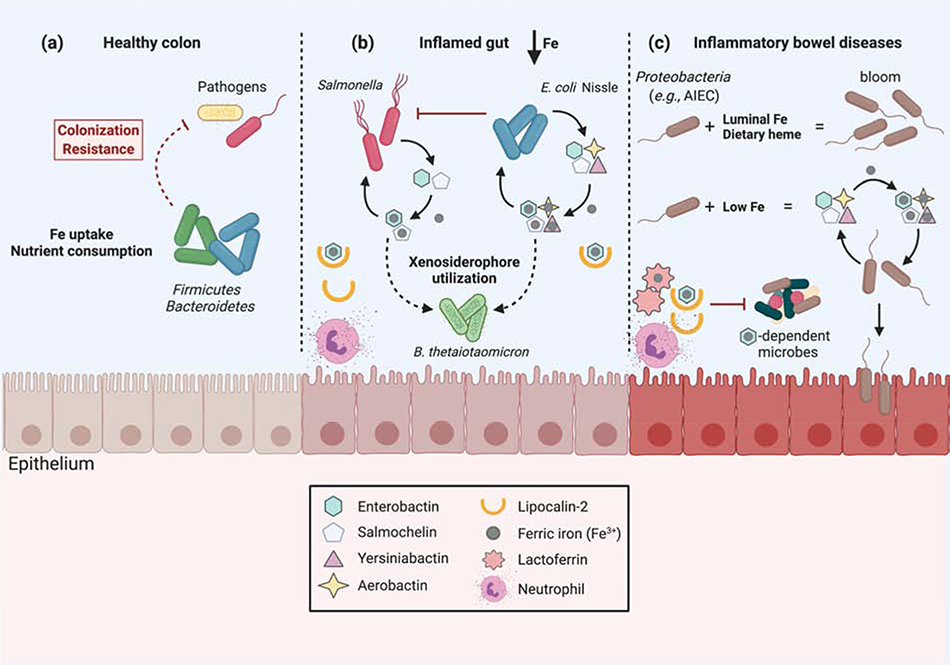
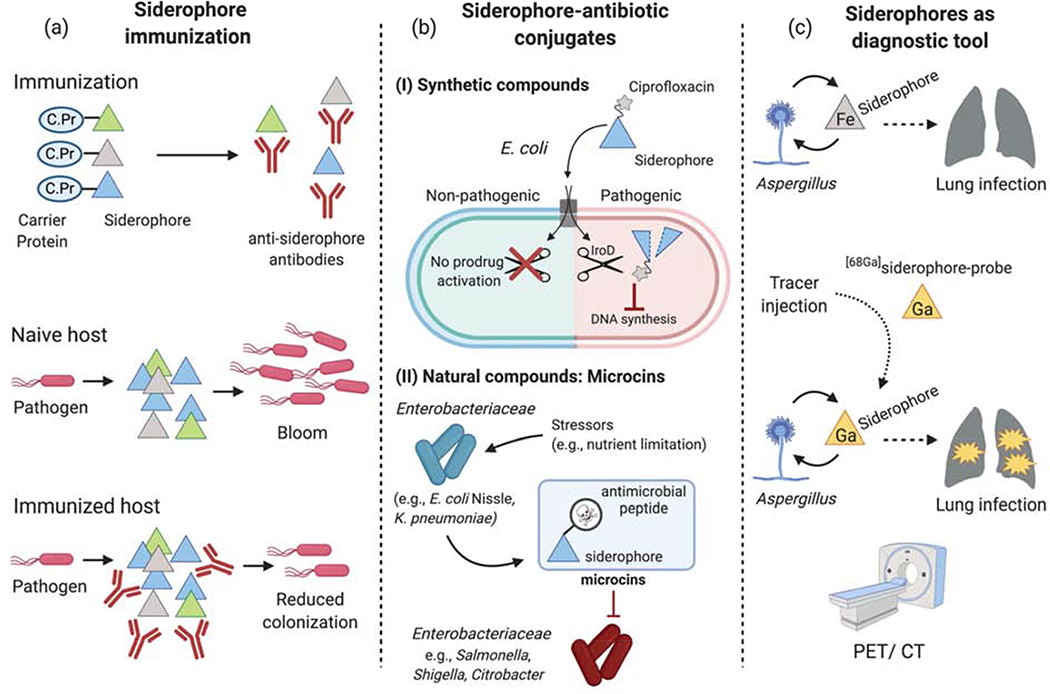
References
-
- Almeida RS, Wilson D, and Hube B (2009). Candida albicans iron acquisition within the host. FEMS Yeast Res. 9, 1000–1012. - PubMed
-
- Archibald F (1983). Lactobacillus plantarum, an organism not requiring iron. FEMS Microbiol. Lett. 19, 29–32.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
