

HtsRC-Mediated Accumulation of F-Actin Regulates Ring Canal Size During Drosophila melanogaster Oogenesis
- PMID: 32883702
- PMCID: PMC7648574
- DOI: 10.1534/genetics.120.303629
HtsRC-Mediated Accumulation of F-Actin Regulates Ring Canal Size During Drosophila melanogaster Oogenesis
Abstract
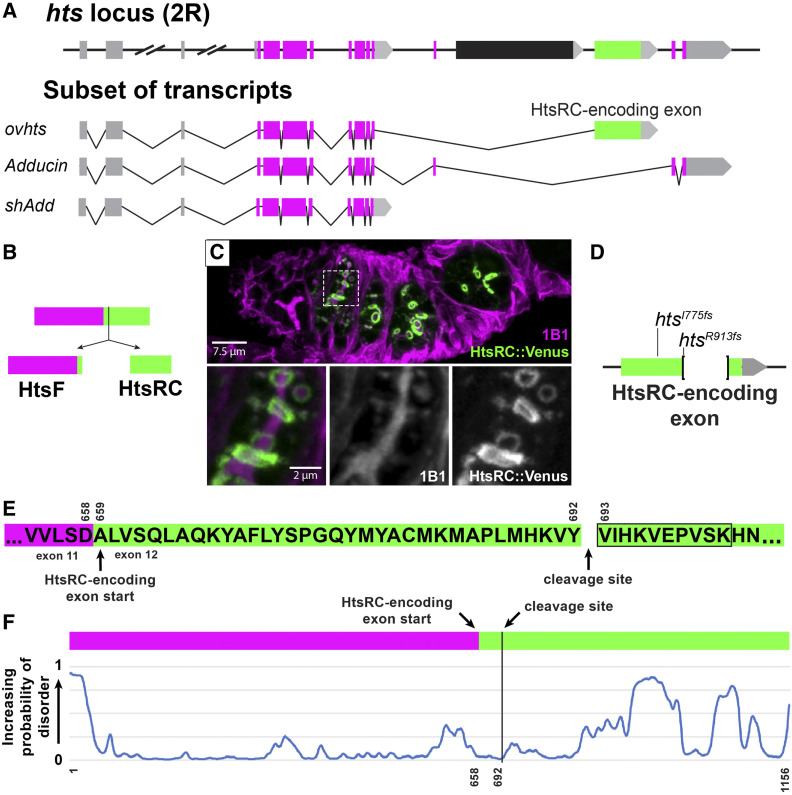
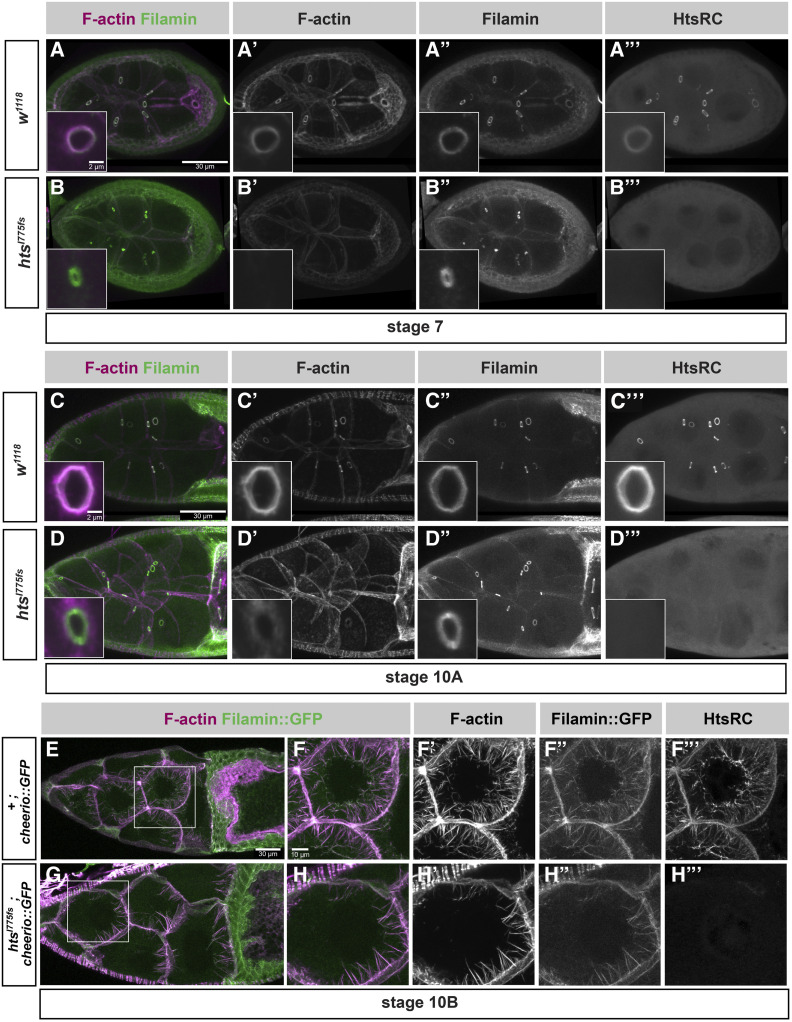
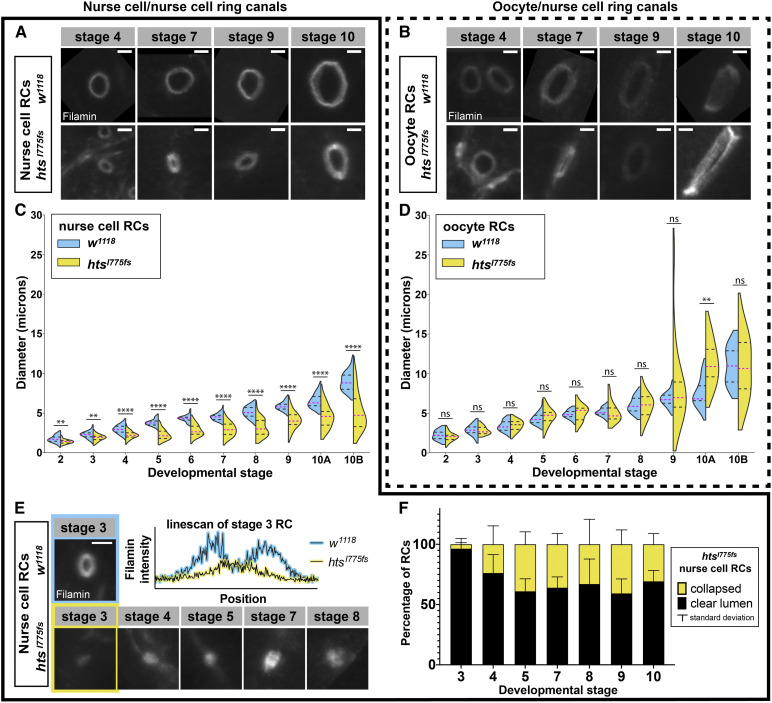
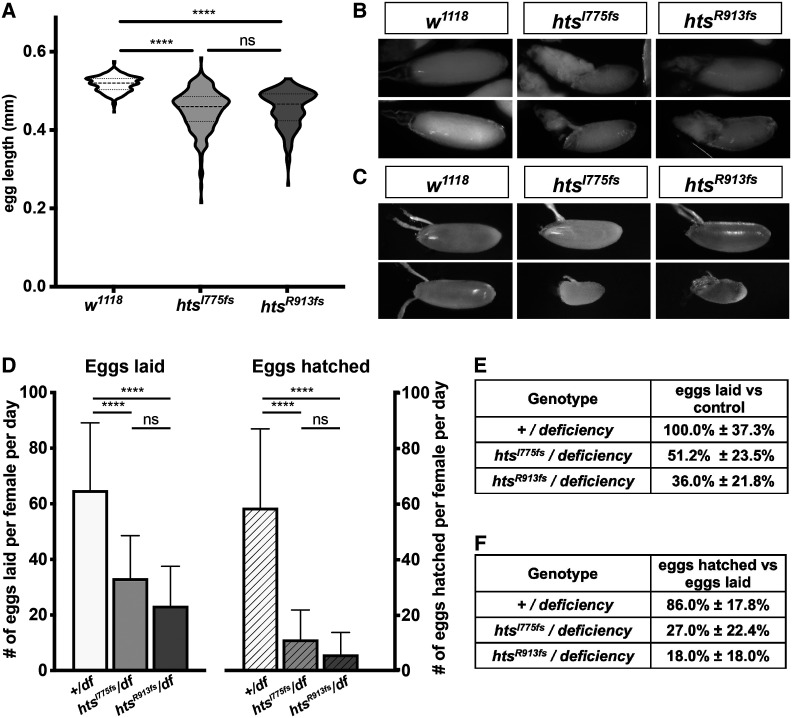
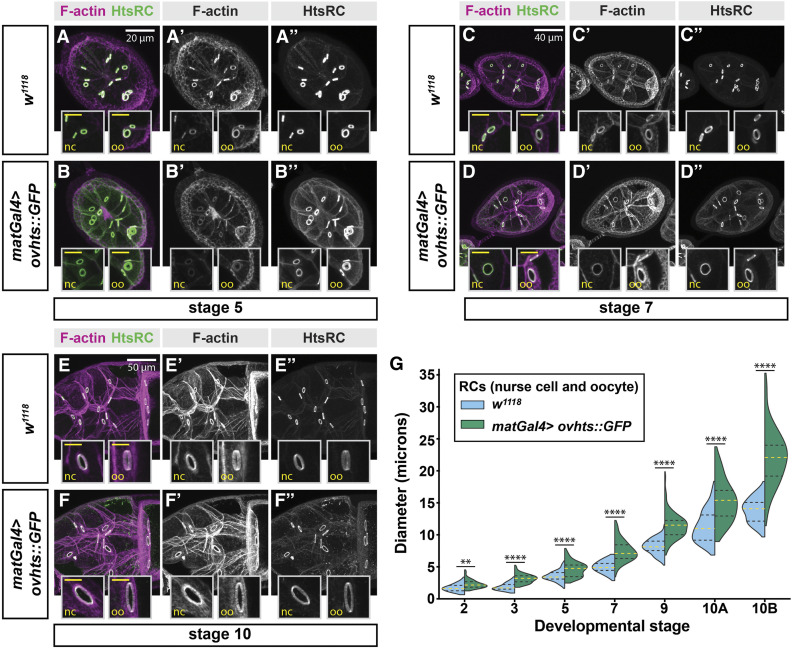
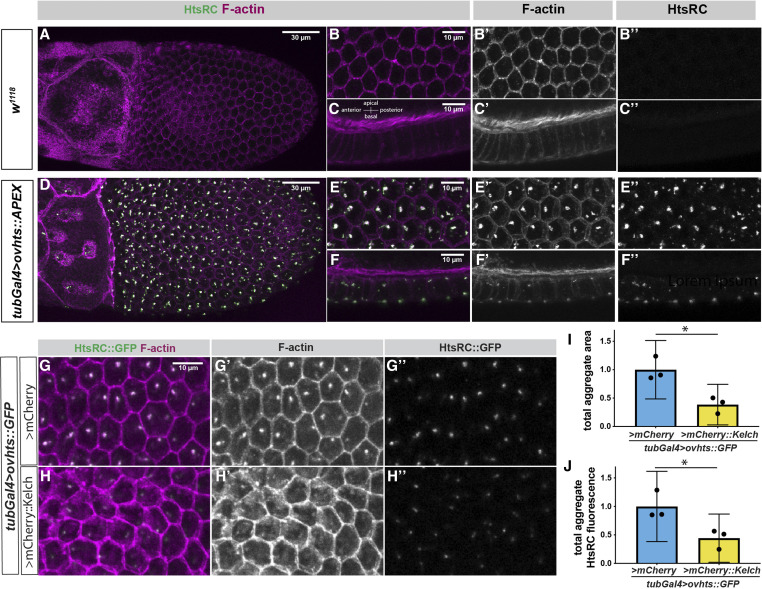
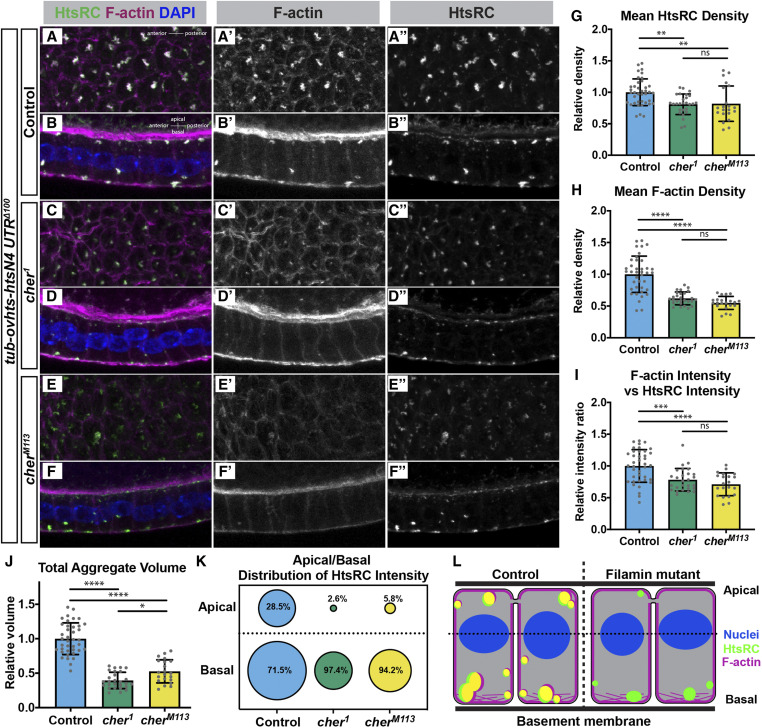
Ring canals in the female germline of Drosophila melanogaster are supported by a robust filamentous actin (F-actin) cytoskeleton, setting them apart from ring canals in other species and tissues. Previous work has identified components required for the expansion of the ring canal actin cytoskeleton, but has not identified the proteins responsible for F-actin recruitment or accumulation. Using a combination of CRISPR-Cas9 mediated mutagenesis and UAS-Gal4 overexpression, we show that HtsRC-a component specific to female germline ring canals-is both necessary and sufficient to drive F-actin accumulation. Absence of HtsRC in the germline resulted in ring canals lacking inner rim F-actin, while overexpression of HtsRC led to larger ring canals. HtsRC functions in combination with Filamin to recruit F-actin to ectopic actin structures in somatic follicle cells. Finally, we present findings that indicate that HtsRC expression and robust female germline ring canal expansion are important for high fecundity in fruit flies but dispensable for their fertility-a result that is consistent with our understanding of HtsRC as a newly evolved gene specific to female germline ring canals.
Keywords: Drosophila oogenesis; actin cytoskeleton; hu li tai shao; intrinsically disordered protein; ring canal.
Copyright © 2020 by the Genetics Society of America.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
