

Structure of a human 48 S translational initiation complex
- PMID: 32883864
- PMCID: PMC7116333
- DOI: 10.1126/science.aba4904
Structure of a human 48 S translational initiation complex
Abstract
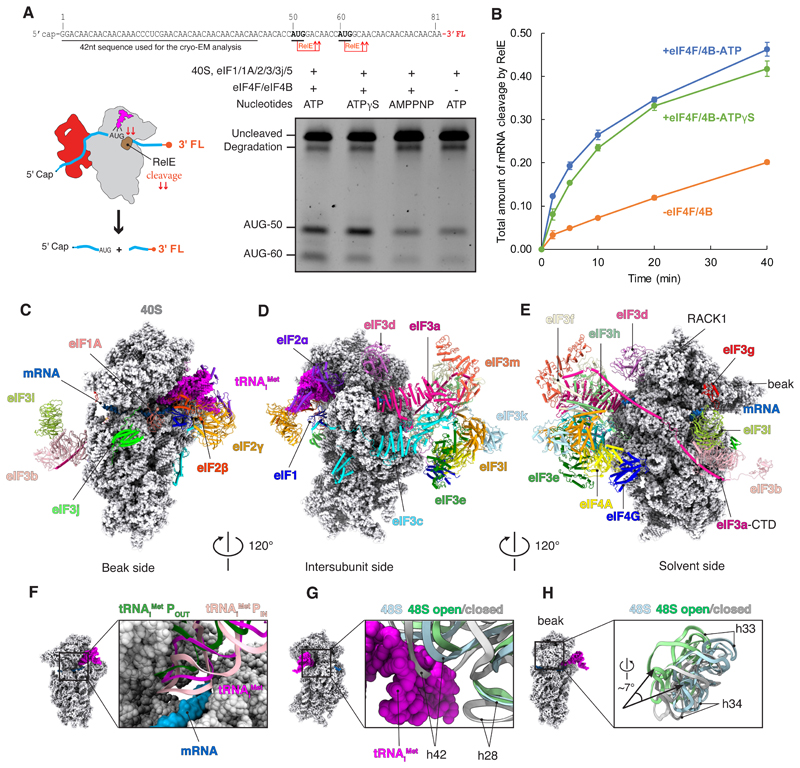
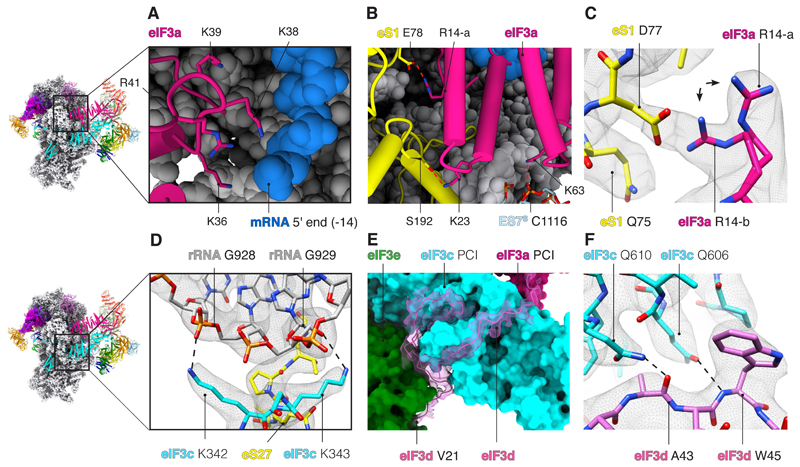
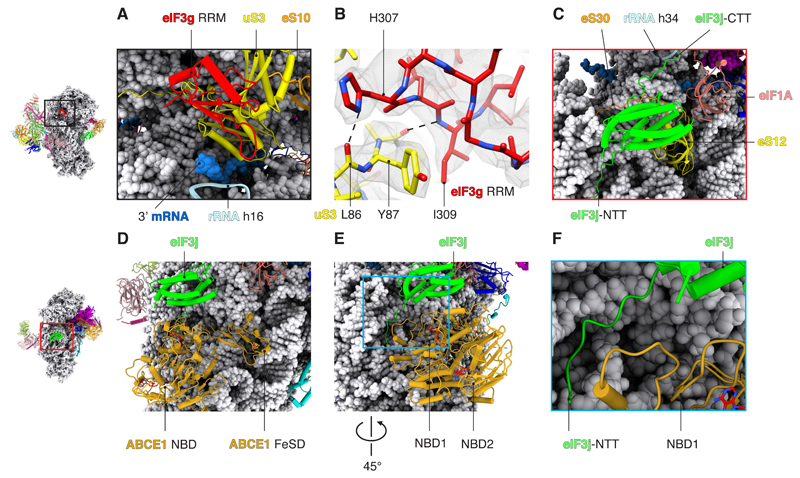
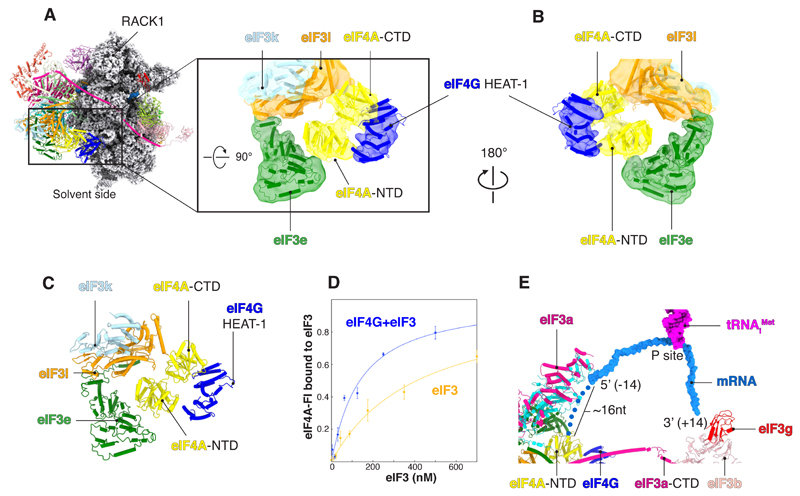
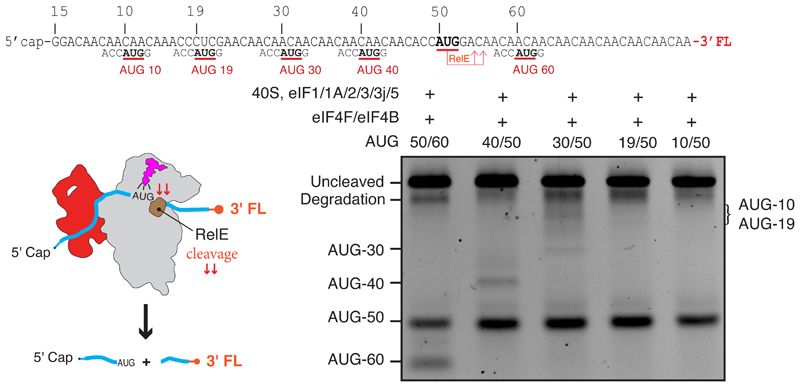
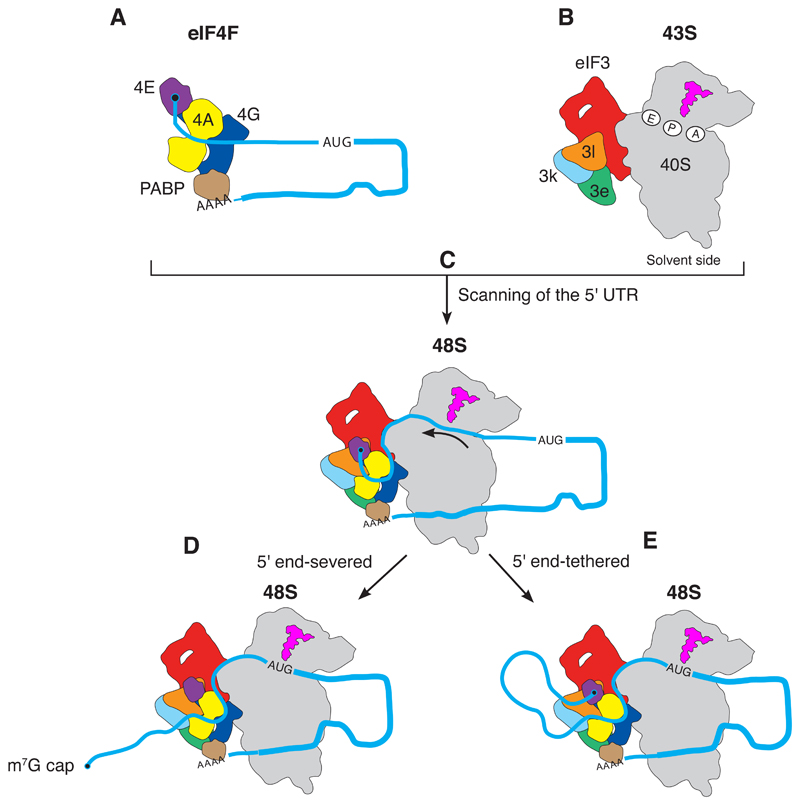
A key step in translational initiation is the recruitment of the 43S preinitiation complex by the cap-binding complex [eukaryotic initiation factor 4F (eIF4F)] at the 5' end of messenger RNA (mRNA) to form the 48S initiation complex (i.e., the 48S). The 48S then scans along the mRNA to locate a start codon. To understand the mechanisms involved, we used cryo-electron microscopy to determine the structure of a reconstituted human 48S The structure reveals insights into early events of translation initiation complex assembly, as well as how eIF4F interacts with subunits of eIF3 near the mRNA exit channel in the 43S The location of eIF4F is consistent with a slotting model of mRNA recruitment and suggests that downstream mRNA is unwound at least in part by being "pulled" through the 40S subunit during scanning.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
-
- Grifo JA, Tahara SM, Morgan MA, Shatkin AJ, Merrick WC. New initiation factor activity required for globin mRNA translation. J Biol Chem. 1983;258:5804–5810. Medline. - PubMed
-
- Morino S, Imataka H, Svitkin YV, Pestova TV, Sonenberg N. Eukaryotic translation initiation factor 4E (eIF4E) binding site and the middle one-third of eIF4GI constitute the core domain for cap-dependent translation, and the C-terminal one-third functions as a modulatory region. Mol Cell Biol. 2000;20:468–477. doi: 10.1128/MCB.20.2.468-477.2000. Medline. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
