

A phylogenetic view and functional annotation of the animal β1,3-glycosyltransferases of the GT31 CAZy family
- PMID: 32886776
- PMCID: PMC8022947
- DOI: 10.1093/glycob/cwaa086
A phylogenetic view and functional annotation of the animal β1,3-glycosyltransferases of the GT31 CAZy family
Abstract
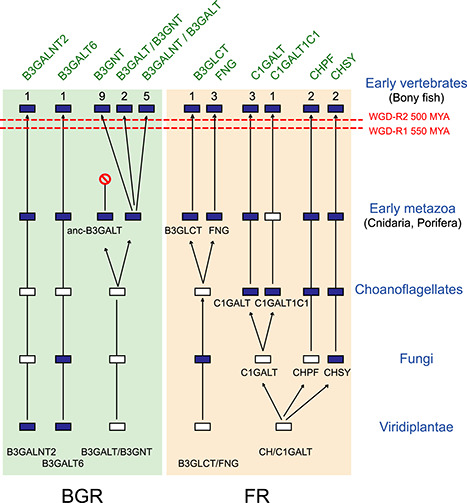
The formation of β1,3-linkages on animal glycoconjugates is catalyzed by a subset of β1,3-glycosyltransferases grouped in the Carbohydrate-Active enZYmes family glycosyltransferase-31 (GT31). This family represents an extremely diverse set of β1,3-N-acetylglucosaminyltransferases [B3GNTs and Fringe β1,3-N-acetylglucosaminyltransferases], β1,3-N-acetylgalactosaminyltransferases (B3GALNTs), β1,3-galactosyltransferases [B3GALTs and core 1 β1,3-galactosyltransferases (C1GALTs)], β1,3-glucosyltransferase (B3GLCT) and β1,3-glucuronyl acid transferases (B3GLCATs or CHs). The mammalian enzymes were particularly well studied and shown to use a large variety of sugar donors and acceptor substrates leading to the formation of β1,3-linkages in various glycosylation pathways. In contrast, there are only a few studies related to other metazoan and lower vertebrates GT31 enzymes and the evolutionary relationships of these divergent sequences remain obscure. In this study, we used bioinformatics approaches to identify more than 920 of putative GT31 sequences in Metazoa, Fungi and Choanoflagellata revealing their deep ancestry. Sequence-based analysis shed light on conserved motifs and structural features that are signatures of all the GT31. We leverage pieces of evidence from gene structure, phylogenetic and sequence-based analyses to identify two major subgroups of GT31 named Fringe-related and B3GALT-related and demonstrate the existence of 10 orthologue groups in the Urmetazoa, the hypothetical last common ancestor of all animals. Finally, synteny and paralogy analysis unveiled the existence of 30 subfamilies in vertebrates, among which 5 are new and were named C1GALT2, C1GALT3, B3GALT8, B3GNT10 and B3GNT11. Altogether, these various approaches enabled us to propose the first comprehensive analysis of the metazoan GT31 disentangling their evolutionary relationships.
Keywords: evolution; functional genomics; molecular phylogeny; motifs; β1,3-glycosyltransferases.
© The Author(s) 2020. Published by Oxford University Press.
Figures







References
-
- Amado M, Almeida R, Carneiro F, Levery SB, Holmes EH, Nomoto M, Hollingsworth MA, Hassan H, Schwientek T, Nielsen PA et al. 1998. A family of human β3-galactosyltransferases: Characterization of four members of a UDP-galactose:β-N-acetyl-glucosamine/β-N-acetyl-galactosamine β-1,3-galactosyltransferase family. J Biol Chem. 273:12770–12778. - PubMed
-
- Amores A, Force A, Yan YL, Joly L, Amemiya C, Fritz A, Ho RK, Langeland J, Prince V, Wang YL et al. 1998. Zebrafish hox clusters and vertebrate genome evolution. Science. 282:1711–1714. - PubMed
