

Dystonia genes functionally converge in specific neurons and share neurobiology with psychiatric disorders
- PMID: 32889528
- PMCID: PMC8354373
- DOI: 10.1093/brain/awaa217
Dystonia genes functionally converge in specific neurons and share neurobiology with psychiatric disorders
Erratum in
-
Erratum to: Dystonia genes functionally converge in specific neurons and share neurobiology with psychiatric disorders.Brain. 2021 Feb 12;144(1):e13. doi: 10.1093/brain/awaa366. Brain. 2021. PMID: 33578419 Free PMC article. No abstract available.
Abstract
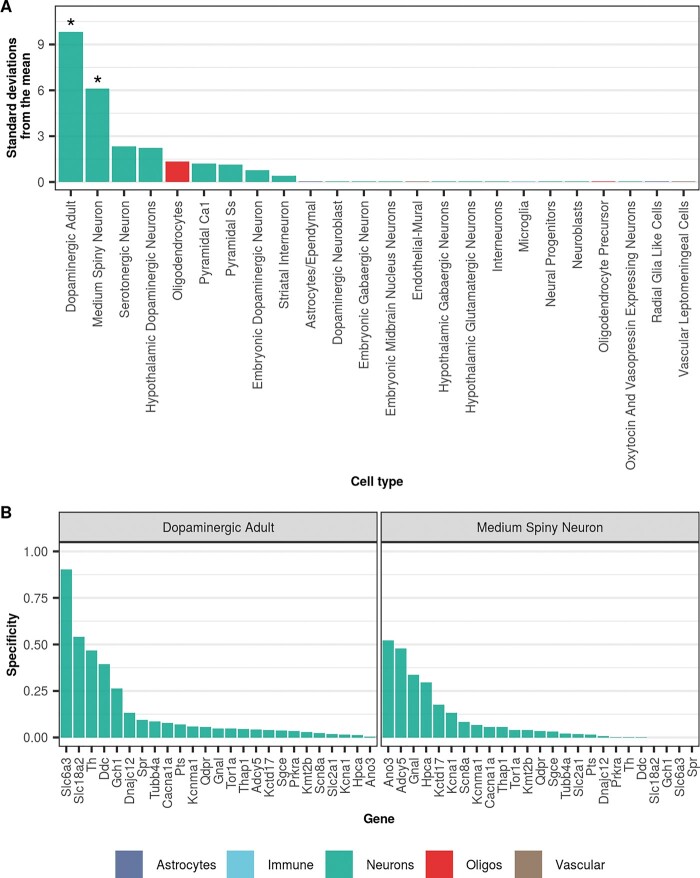
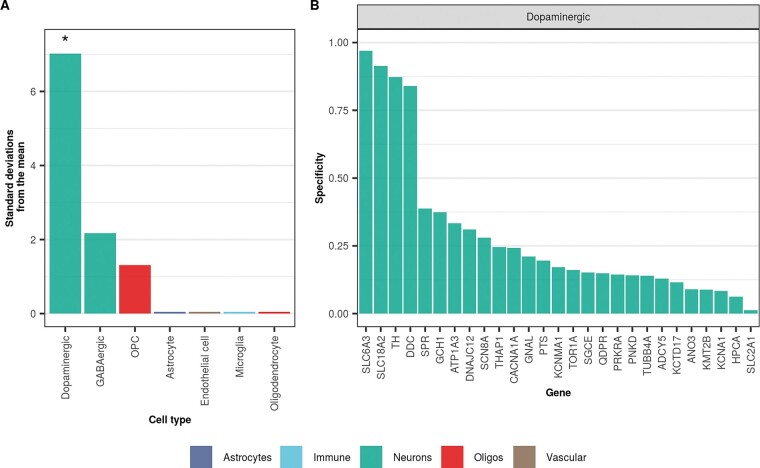
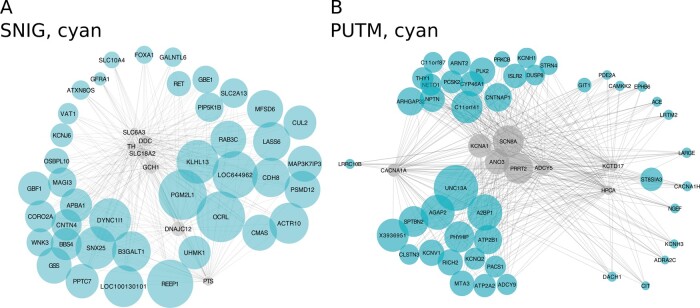
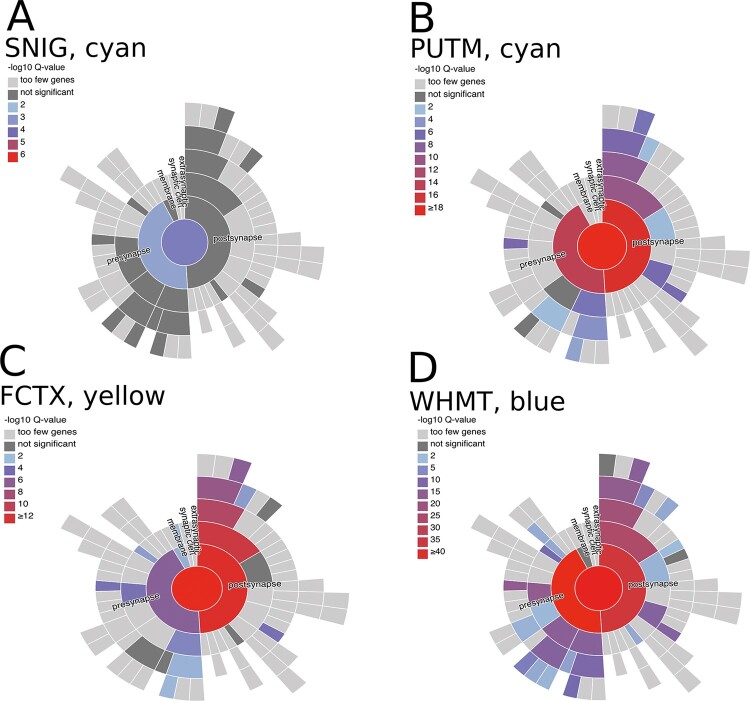
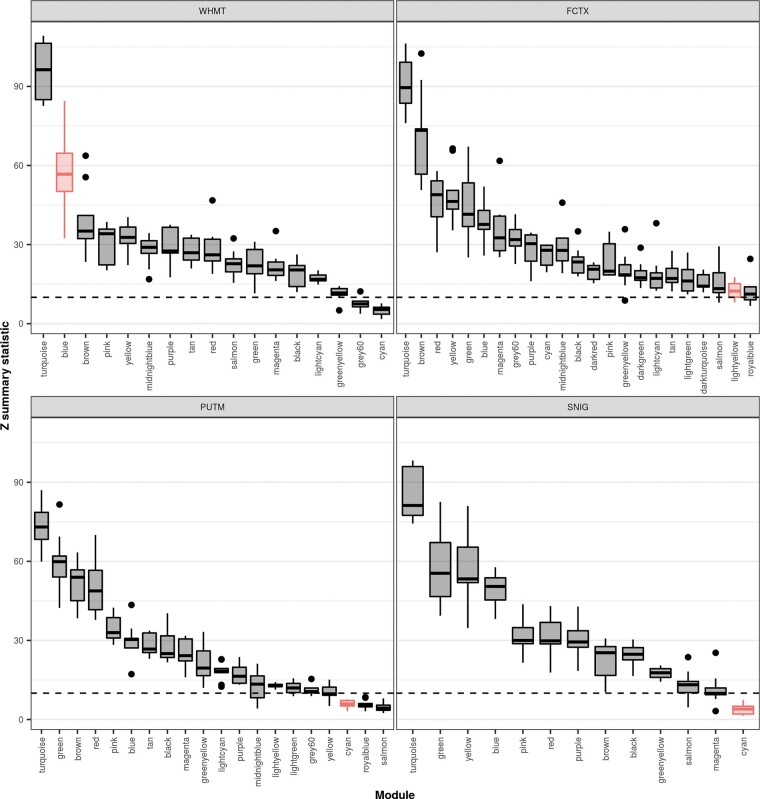
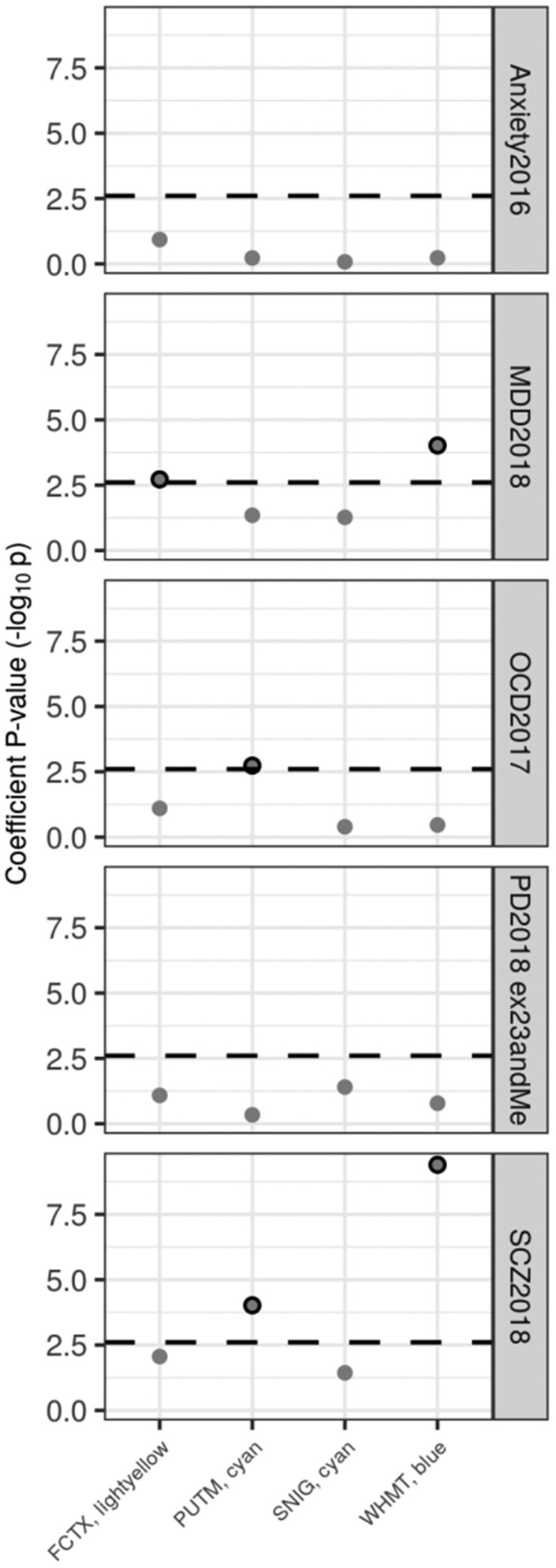
Dystonia is a neurological disorder characterized by sustained or intermittent muscle contractions causing abnormal movements and postures, often occurring in absence of any structural brain abnormality. Psychiatric comorbidities, including anxiety, depression, obsessive-compulsive disorder and schizophrenia, are frequent in patients with dystonia. While mutations in a fast-growing number of genes have been linked to Mendelian forms of dystonia, the cellular, anatomical, and molecular basis remains unknown for most genetic forms of dystonia, as does its genetic and biological relationship to neuropsychiatric disorders. Here we applied an unbiased systems-biology approach to explore the cellular specificity of all currently known dystonia-associated genes, predict their functional relationships, and test whether dystonia and neuropsychiatric disorders share a genetic relationship. To determine the cellular specificity of dystonia-associated genes in the brain, single-nuclear transcriptomic data derived from mouse brain was used together with expression-weighted cell-type enrichment. To identify functional relationships among dystonia-associated genes, we determined the enrichment of these genes in co-expression networks constructed from 10 human brain regions. Stratified linkage-disequilibrium score regression was used to test whether co-expression modules enriched for dystonia-associated genes significantly contribute to the heritability of anxiety, major depressive disorder, obsessive-compulsive disorder, schizophrenia, and Parkinson's disease. Dystonia-associated genes were significantly enriched in adult nigral dopaminergic neurons and striatal medium spiny neurons. Furthermore, 4 of 220 gene co-expression modules tested were significantly enriched for the dystonia-associated genes. The identified modules were derived from the substantia nigra, putamen, frontal cortex, and white matter, and were all significantly enriched for genes associated with synaptic function. Finally, we demonstrate significant enrichments of the heritability of major depressive disorder, obsessive-compulsive disorder and schizophrenia within the putamen and white matter modules, and a significant enrichment of the heritability of Parkinson's disease within the substantia nigra module. In conclusion, multiple dystonia-associated genes interact and contribute to pathogenesis likely through dysregulation of synaptic signalling in striatal medium spiny neurons, adult nigral dopaminergic neurons and frontal cortical neurons. Furthermore, the enrichment of the heritability of psychiatric disorders in the co-expression modules enriched for dystonia-associated genes indicates that psychiatric symptoms associated with dystonia are likely to be intrinsic to its pathophysiology.
Keywords: dystonia; medium-spiny neurons; network analysis; synaptic transmission; transcriptomic analysis.
© The Author(s) (2020). Published by Oxford University Press on behalf of the Guarantors of Brain.
Figures
Comment in
-
'Moving genes': how dystonia genes functionally converge on the transcriptome.Brain. 2020 Sep 1;143(9):2631-2634. doi: 10.1093/brain/awaa253. Brain. 2020. PMID: 32947615 No abstract available.
-
Neurology and Psychiatry Get Closer Again: Lessons From Dystonia.Mov Disord. 2021 Jan;36(1):85-86. doi: 10.1002/mds.28414. Epub 2020 Nov 30. Mov Disord. 2021. PMID: 33253434 No abstract available.
References
-
- Atasu B, Hanagasi H, Bilgic B, Pak M, Erginel-Unaltuna N, Hauser AK, et al. HPCA confirmed as a genetic cause of DYT2-like dystonia phenotype. Mov Disord 2018; 33: 1354–8. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
