

A long non-coding RNA (Lrap) modulates brain gene expression and levels of alcohol consumption in rats
- PMID: 32893479
- PMCID: PMC7900948
- DOI: 10.1111/gbb.12698
A long non-coding RNA (Lrap) modulates brain gene expression and levels of alcohol consumption in rats
Abstract
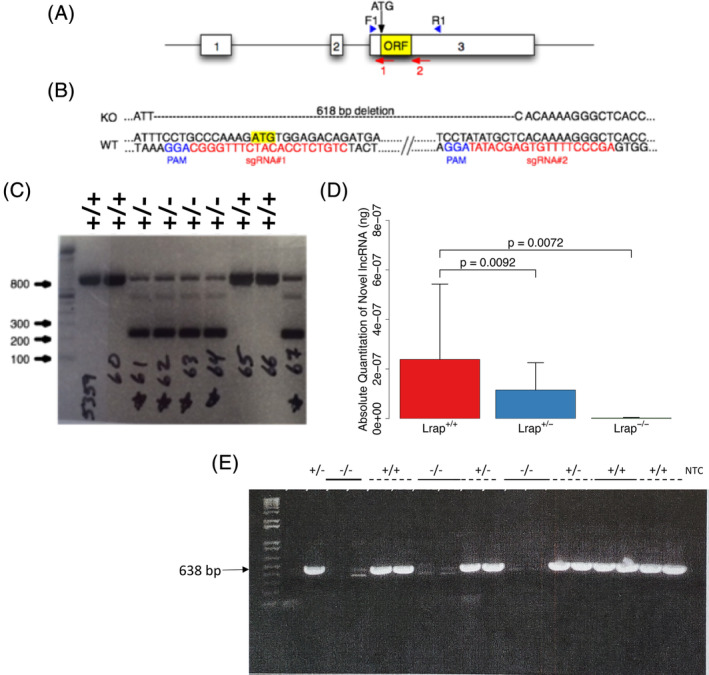
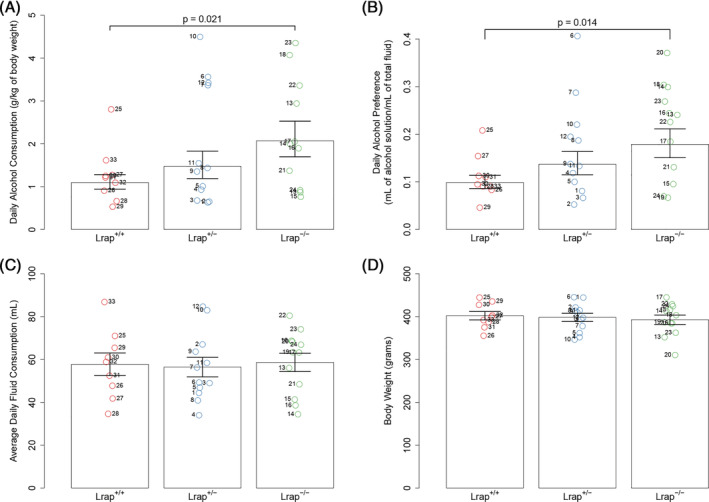
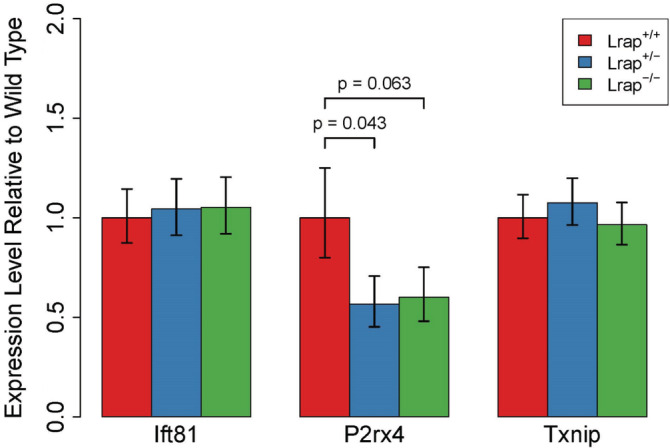
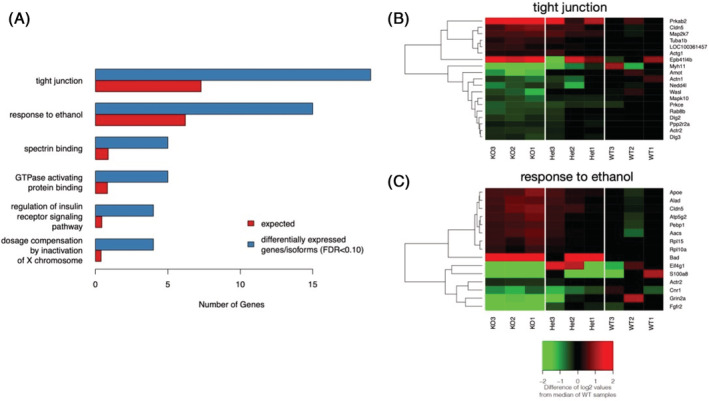
LncRNAs are important regulators of quantitative and qualitative features of the transcriptome. We have used QTL and other statistical analyses to identify a gene coexpression module associated with alcohol consumption. The "hub gene" of this module, Lrap (Long non-coding RNA for alcohol preference), was an unannotated transcript resembling a lncRNA. We used partial correlation analyses to establish that Lrap is a major contributor to the integrity of the coexpression module. Using CRISPR/Cas9 technology, we disrupted an exon of Lrap in Wistar rats. Measures of alcohol consumption in wild type, heterozygous and knockout rats showed that disruption of Lrap produced increases in alcohol consumption/alcohol preference. The disruption of Lrap also produced changes in expression of over 700 other transcripts. Furthermore, it became apparent that Lrap may have a function in alternative splicing of the affected transcripts. The GO category of "Response to Ethanol" emerged as one of the top candidates in an enrichment analysis of the differentially expressed transcripts. We validate the role of Lrap as a mediator of alcohol consumption by rats, and also implicate Lrap as a modifier of the expression and splicing of a large number of brain transcripts. A defined subset of these transcripts significantly impacts alcohol consumption by rats (and possibly humans). Our work shows the pleiotropic nature of non-coding elements of the genome, the power of network analysis in identifying the critical elements influencing phenotypes, and the fact that not all changes produced by genetic editing are critical for the concomitant changes in phenotype.
Keywords: CRISPR/Cas; brain RNA expression networks; genetic modification; long non-coding RNA; predisposing factors; quantitative genetics; recombinant inbred rat strains; systems genetics; transcriptome; voluntary alcohol consumption.
© 2020 The Authors. Genes, Brain and Behavior published by International Behavioural and Neural Genetics Society and John Wiley & Sons Ltd.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
