

Network Rewiring: Physiological Consequences of Reciprocally Exchanging the Physical Locations and Growth-Phase-Dependent Expression Patterns of the Salmonella fis and dps Genes
- PMID: 32900812
- PMCID: PMC7482072
- DOI: 10.1128/mBio.02128-20
Network Rewiring: Physiological Consequences of Reciprocally Exchanging the Physical Locations and Growth-Phase-Dependent Expression Patterns of the Salmonella fis and dps Genes
Abstract
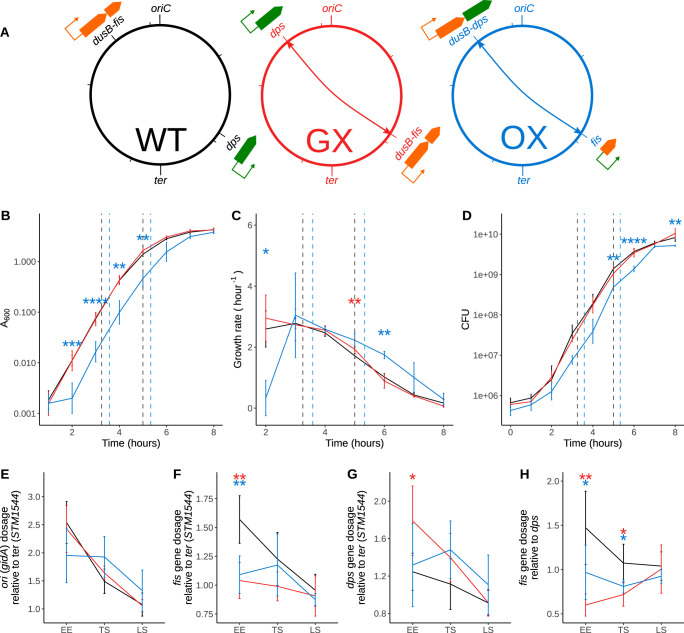
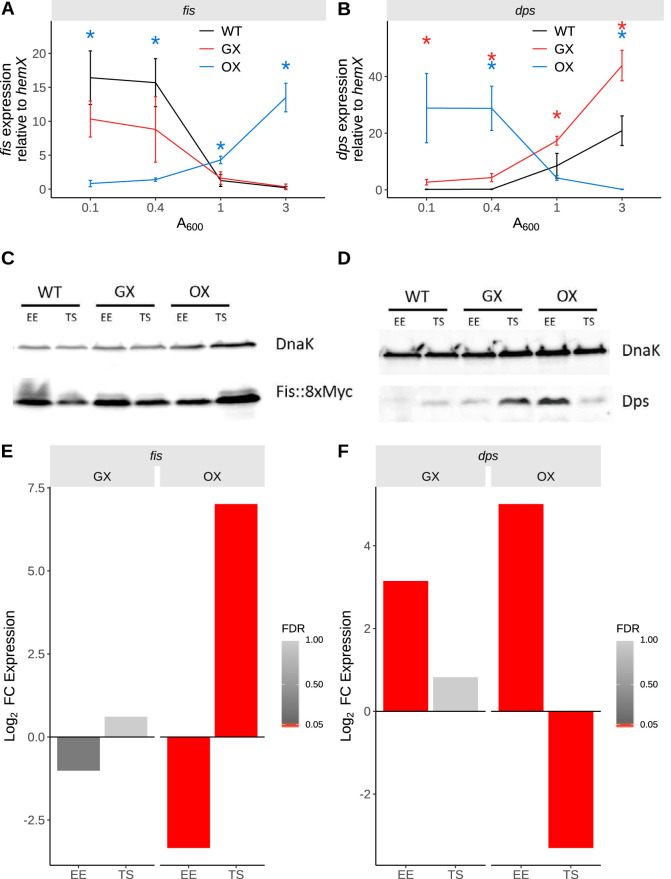
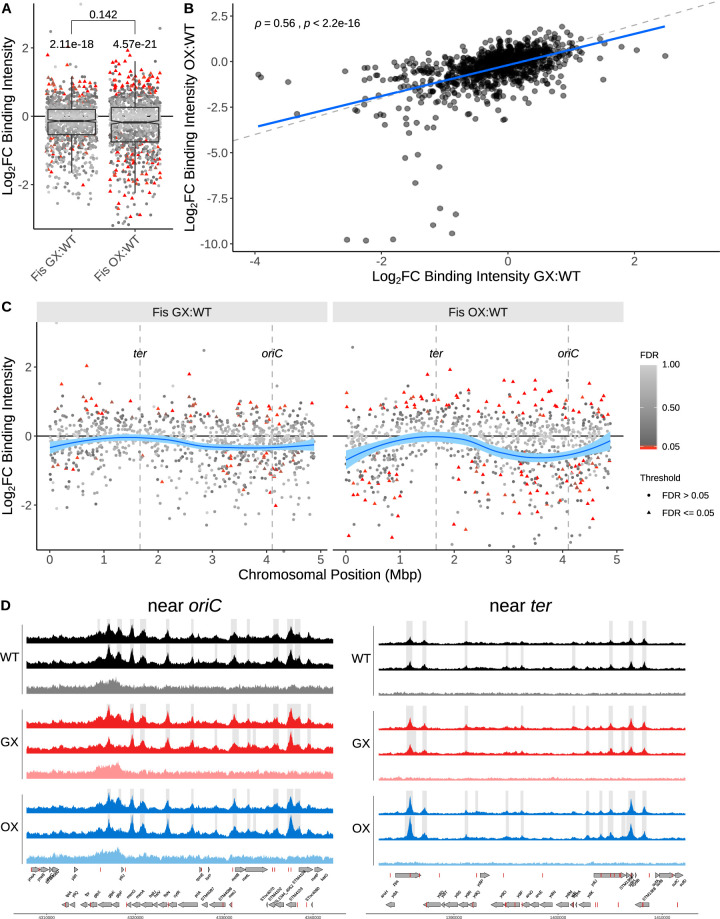
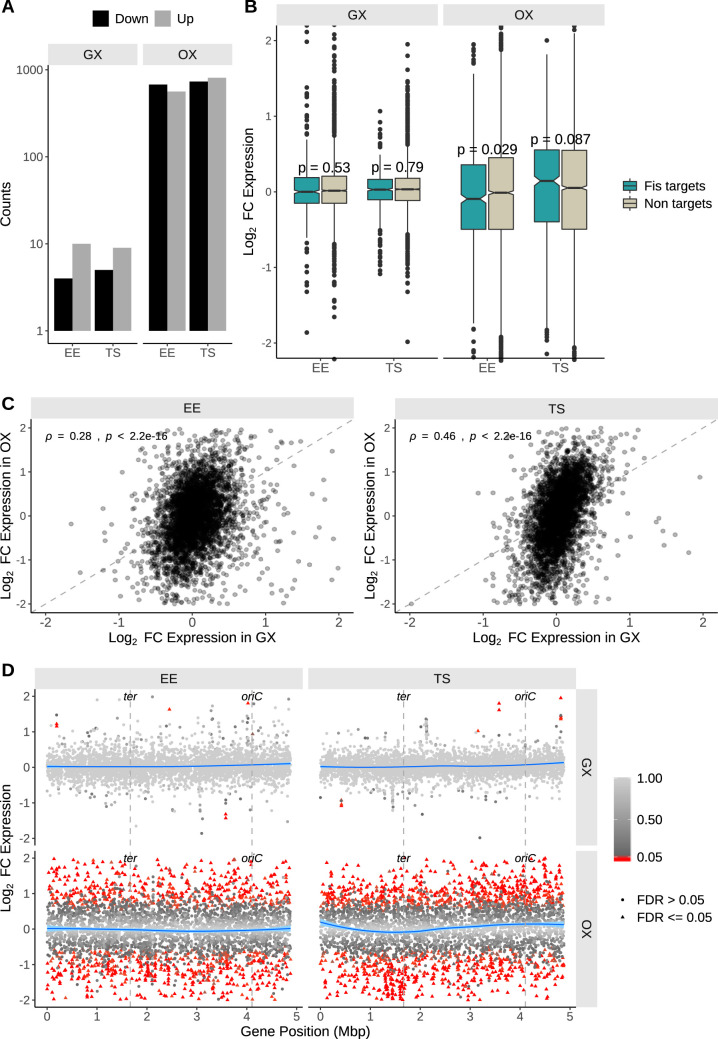
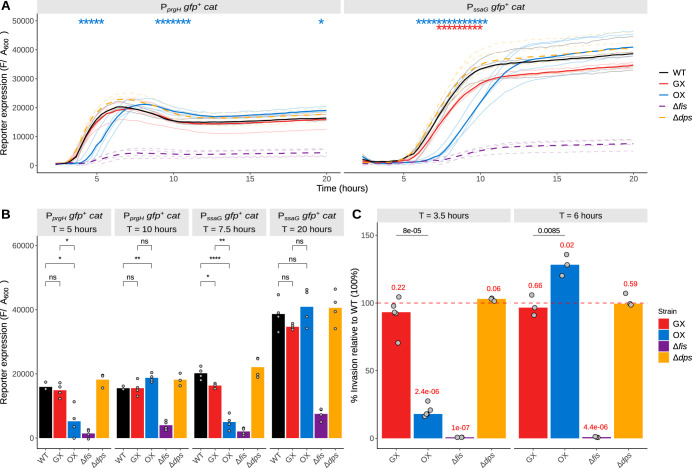
The Fis nucleoid-associated protein controls the expression of a large and diverse regulon of genes in Gram-negative bacteria. Fis production is normally maximal in bacteria during the early exponential phase of batch culture growth, becoming almost undetectable by the onset of stationary phase. We tested the effect on the Fis regulatory network in Salmonella of moving the complete fis gene from its usual location near the origin of chromosomal replication to the position normally occupied by the dps gene in the right macrodomain of the chromosome, and vice versa, creating the
Keywords: ChIP-seq; Dps; Fis; RNA-seq; Salmonella enterica serovar Typhimurium; nucleoid-associated protein; transcriptome; virulence.
Copyright © 2020 Bogue et al.
Figures
Similar articles
-
Reciprocally rewiring and repositioning the Integration Host Factor (IHF) subunit genes in Salmonella enterica serovar Typhimurium: impacts on physiology and virulence.Microb Genom. 2022 Feb;8(2):000768. doi: 10.1099/mgen.0.000768. Microb Genom. 2022. PMID: 35166652 Free PMC article.
-
Genome-wide analysis of the salmonella Fis regulon and its regulatory mechanism on pathogenicity islands.PLoS One. 2013 May 23;8(5):e64688. doi: 10.1371/journal.pone.0064688. Print 2013. PLoS One. 2013. PMID: 23717649 Free PMC article.
-
Expression of the Fis protein is sustained in late-exponential- and stationary-phase cultures of Salmonella enterica serovar Typhimurium grown in the absence of aeration.Mol Microbiol. 2007 Oct;66(1):237-51. doi: 10.1111/j.1365-2958.2007.05916.x. Epub 2007 Sep 3. Mol Microbiol. 2007. PMID: 17784910
-
Physiological Robustness of Model Gram-Negative Bacteria in Response to Genome Rewiring.Microb Physiol. 2022;32(5-6):158-176. doi: 10.1159/000526651. Epub 2022 Aug 30. Microb Physiol. 2022. PMID: 36041419 Review.
-
Bacterial virulence and Fis: adapting regulatory networks to the host environment.Trends Microbiol. 2014 Feb;22(2):92-9. doi: 10.1016/j.tim.2013.11.008. Epub 2013 Dec 24. Trends Microbiol. 2014. PMID: 24370464 Review.
Cited by
-
Nucleoid-associated proteins shape chromatin structure and transcriptional regulation across the bacterial kingdom.Transcription. 2021 Aug;12(4):182-218. doi: 10.1080/21541264.2021.1973865. Epub 2021 Sep 9. Transcription. 2021. PMID: 34499567 Free PMC article. Review.
-
Composition of Transcription Machinery and Its Crosstalk with Nucleoid-Associated Proteins and Global Transcription Factors.Biomolecules. 2021 Jun 22;11(7):924. doi: 10.3390/biom11070924. Biomolecules. 2021. PMID: 34206477 Free PMC article. Review.
-
The regulation of DNA supercoiling across evolution.Protein Sci. 2021 Oct;30(10):2042-2056. doi: 10.1002/pro.4171. Epub 2021 Aug 23. Protein Sci. 2021. PMID: 34398513 Free PMC article. Review.
-
The DNA relaxation-dependent OFF-to-ON biasing of the type 1 fimbrial genetic switch requires the Fis nucleoid-associated protein.Microbiology (Reading). 2023 Jan;169(1):001283. doi: 10.1099/mic.0.001283. Microbiology (Reading). 2023. PMID: 36748578 Free PMC article.
-
Reciprocally rewiring and repositioning the Integration Host Factor (IHF) subunit genes in Salmonella enterica serovar Typhimurium: impacts on physiology and virulence.Microb Genom. 2022 Feb;8(2):000768. doi: 10.1099/mgen.0.000768. Microb Genom. 2022. PMID: 35166652 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
