

Silence as a way of niche adaptation: mecC-MRSA with variations in the accessory gene regulator (agr) functionality express kaleidoscopic phenotypes
- PMID: 32901059
- PMCID: PMC7479134
- DOI: 10.1038/s41598-020-71640-4
Silence as a way of niche adaptation: mecC-MRSA with variations in the accessory gene regulator (agr) functionality express kaleidoscopic phenotypes
Abstract
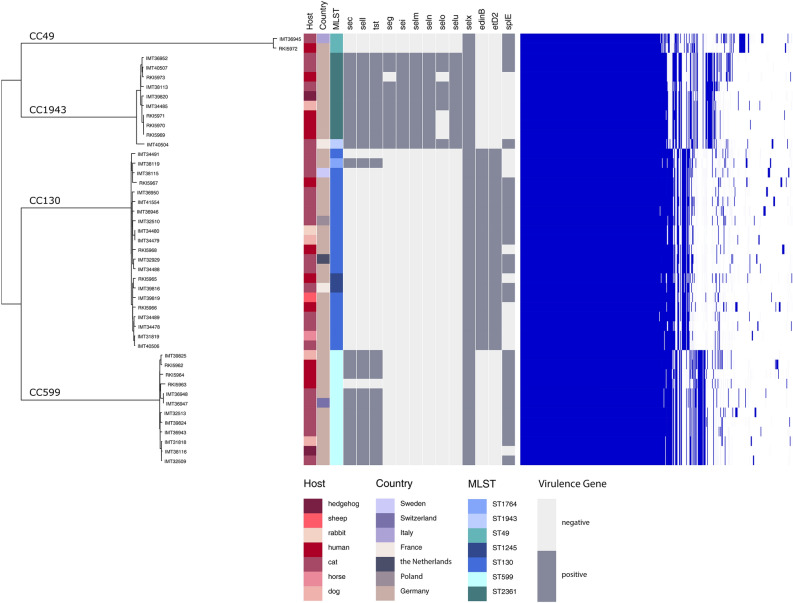
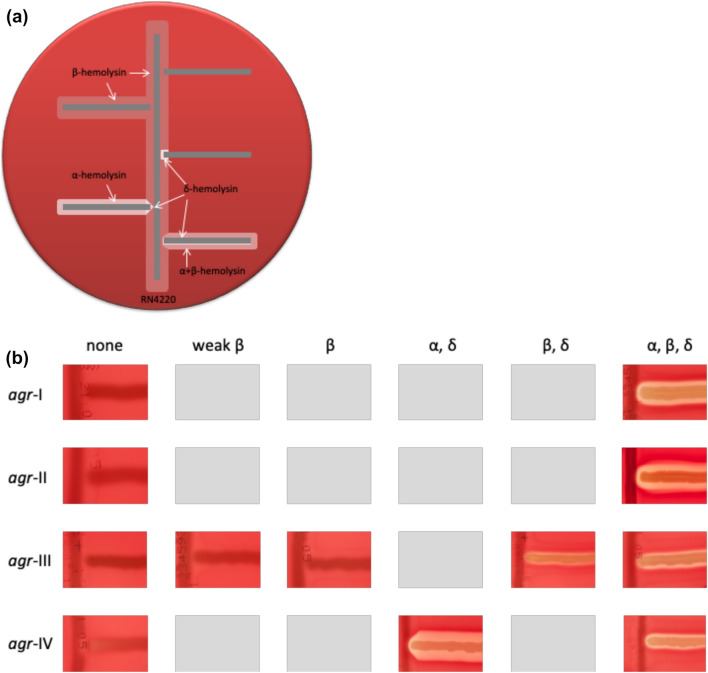
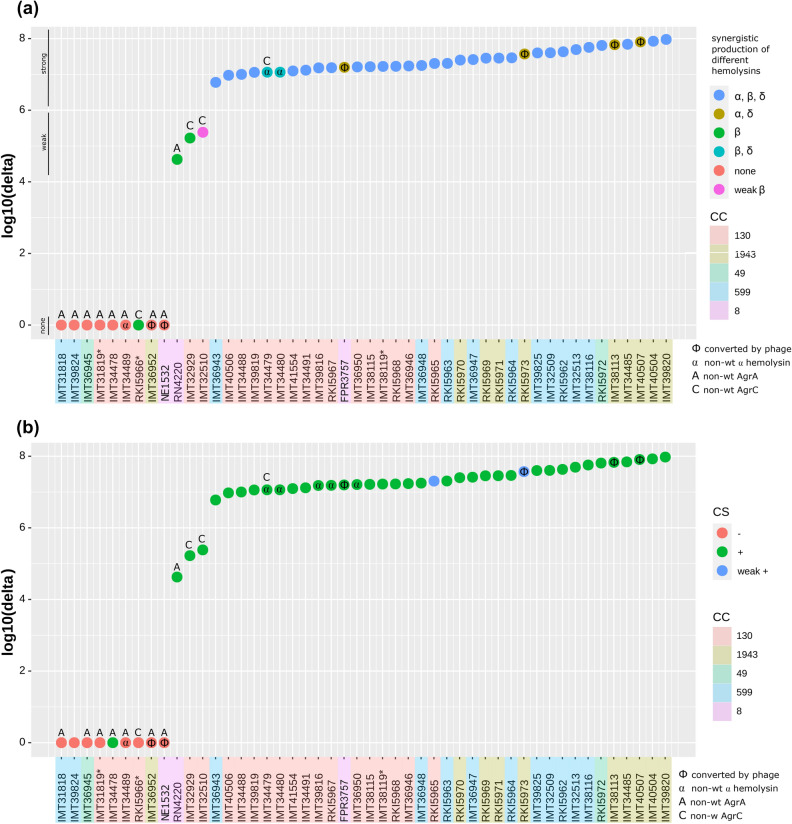
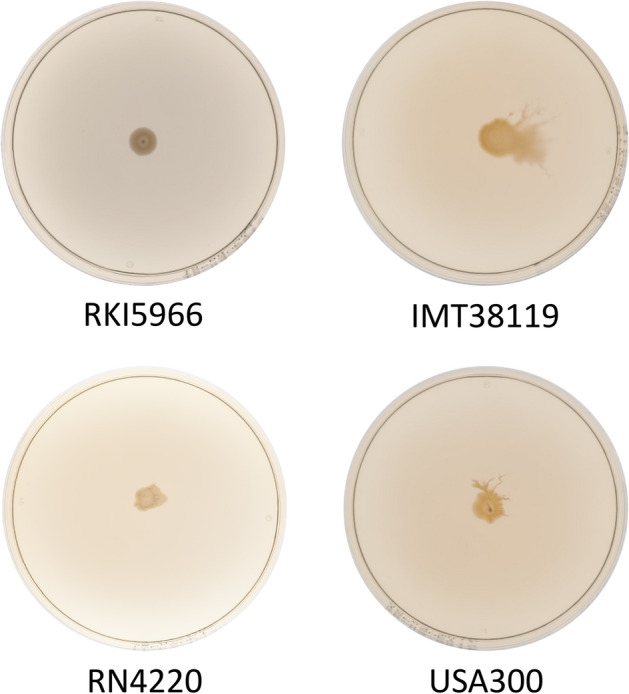
Functionality of the accessory gene regulator (agr) quorum sensing system is an important factor promoting either acute or chronic infections by the notorious opportunistic human and veterinary pathogen Staphylococcus aureus. Spontaneous alterations of the agr system are known to frequently occur in human healthcare-associated S. aureus lineages. However, data on agr integrity and function are sparse regarding other major clonal lineages. Here we report on the agr system functionality and activity level in mecC-carrying methicillin resistant S. aureus (MRSA) of various animal origins (n = 33) obtained in Europe as well as in closely related human isolates (n = 12). Whole genome analysis assigned all isolates to four clonal complexes (CC) with distinct agr types (CC599 agr I, CC49 agr II, CC130 agr III and CC1943 agr IV). Agr functionality was assessed by a combination of phenotypic assays and proteome analysis. In each CC, isolates with varying agr activity levels were detected, including the presence of completely non-functional variants. Genomic comparison of the agr I-IV encoding regions associated these phenotypic differences with variations in the agrA and agrC genes. The genomic changes were detected independently in divergent lineages, suggesting that agr variation might foster viability and adaptation of emerging MRSA lineages to distinct ecological niches.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Quorum Sensing, Virulence, and Antibiotic Resistance of USA100 Methicillin-Resistant Staphylococcus aureus Isolates.mSphere. 2019 Aug 14;4(4):e00553-19. doi: 10.1128/mSphere.00553-19. mSphere. 2019. PMID: 31413175 Free PMC article.
-
Impact of accessory gene regulator (agr) dysfunction on vancomycin pharmacodynamics among Canadian community and health-care associated methicillin-resistant Staphylococcus aureus.Ann Clin Microbiol Antimicrob. 2011 May 20;10:20. doi: 10.1186/1476-0711-10-20. Ann Clin Microbiol Antimicrob. 2011. PMID: 21599878 Free PMC article.
-
Variations in amount of TSST-1 produced by clinical methicillin resistant Staphylococcus aureus (MRSA) isolates and allelic variation in accessory gene regulator (agr) locus.BMC Microbiol. 2009 Mar 10;9:52. doi: 10.1186/1471-2180-9-52. BMC Microbiol. 2009. PMID: 19272162 Free PMC article.
-
Methicillin resistance in Staphylococcus isolates: the "mec alphabet" with specific consideration of mecC, a mec homolog associated with zoonotic S. aureus lineages.Int J Med Microbiol. 2014 Oct;304(7):794-804. doi: 10.1016/j.ijmm.2014.06.007. Epub 2014 Jun 28. Int J Med Microbiol. 2014. PMID: 25034857 Review.
-
Quorum sensing-mediated regulation of staphylococcal virulence and antibiotic resistance.Future Microbiol. 2014;9(5):669-81. doi: 10.2217/fmb.14.31. Future Microbiol. 2014. PMID: 24957093 Review.
Cited by
-
Altered genomic methylation promotes Staphylococcus aureus persistence in hospital environment.Nat Commun. 2024 Nov 7;15(1):9619. doi: 10.1038/s41467-024-54033-3. Nat Commun. 2024. PMID: 39511195 Free PMC article.
-
Panton-Valentine Leukocidin-Positive Staphylococcus aureus in Family and Pet Cat.Emerg Infect Dis. 2024 Aug;30(8):1724-1726. doi: 10.3201/eid3008.231255. Emerg Infect Dis. 2024. PMID: 39043433 Free PMC article.
-
Analysis of the Presence of the Virulence and Regulation Genes from Staphylococcus aureus (S. aureus) in Coagulase Negative Staphylococci and the Influence of the Staphylococcal Cross-Talk on Their Functions.Int J Environ Res Public Health. 2023 Mar 15;20(6):5155. doi: 10.3390/ijerph20065155. Int J Environ Res Public Health. 2023. PMID: 36982064 Free PMC article.
-
Gram-positive ESKAPE pathogens in Germany: A comprehensive analysis of occurrence and resistance development in animal, food, and environmental sources.One Health. 2025 Jun 3;20:101099. doi: 10.1016/j.onehlt.2025.101099. eCollection 2025 Jun. One Health. 2025. PMID: 40535975 Free PMC article. Review.
-
Toxic Shock Syndrome Toxin-1 (TSST-1) in Staphylococcus aureus: Prevalence, Molecular Mechanisms, and Public Health Implications.Toxins (Basel). 2025 Jun 24;17(7):323. doi: 10.3390/toxins17070323. Toxins (Basel). 2025. PMID: 40711134 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
