

Sulfhydryl groups as targets of mercury toxicity
- PMID: 32905350
- PMCID: PMC7470069
- DOI: 10.1016/j.ccr.2020.213343
Sulfhydryl groups as targets of mercury toxicity
Abstract
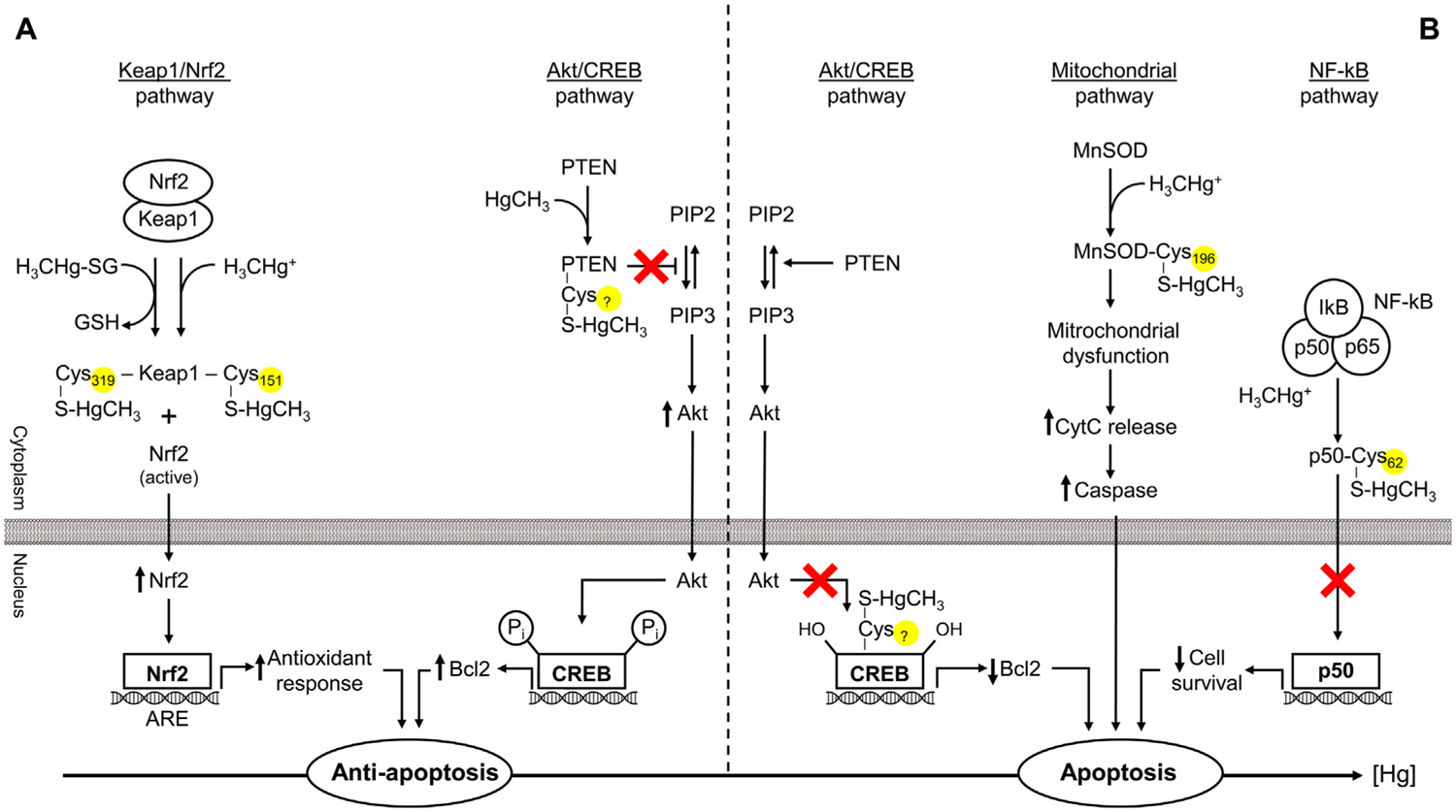
The present study addresses existing data on the affinity and conjugation of sulfhydryl (thiol; -SH) groups of low- and high-molecular-weight biological ligands with mercury (Hg). The consequences of these interactions with special emphasis on pathways of Hg toxicity are highlighted. Cysteine (Cys) is considered the primary target of Hg, and link its sensitivity with thiol groups and cellular damage. In vivo, Hg complexes play a key role in Hg metabolism. Due to the increased affinity of Hg to SH groups in Cys residues, glutathione (GSH) is reactive. The geometry of Hg(II) glutathionates is less understood than that with Cys. Both Cys and GSH Hg-conjugates are important in Hg transport. The binding of Hg to Cys mediates multiple toxic effects of Hg, especially inhibitory effects on enzymes and other proteins that contain free Cys residues. In blood plasma, albumin is the main Hg-binding (Hg2+, CH3Hg+, C2H5Hg+, C6H5Hg+) protein. At the Cys34 residue, Hg2+ binds to albumin, whereas other metals likely are bound at the N-terminal site and multi-metal binding sites. In addition to albumin, Hg binds to multiple Cys-containing enzymes (including manganese-superoxide dismutase (Mn-SOD), arginase I, sorbitol dehydrogenase, and δ-aminolevulinate dehydratase, etc.) involved in multiple processes. The affinity of Hg for thiol groups may also underlie the pathways of Hg toxicity. In particular, Hg-SH may contribute to apoptosis modulation by interfering with Akt/CREB, Keap1/Nrf2, NF-κB, and mitochondrial pathways. Mercury-induced oxidative stress may ensue from Cys-Hg binding and inhibition of Mn-SOD (Cys196), thioredoxin reductase (TrxR) (Cys497) activity, as well as limiting GSH (GS-HgCH3) and Trx (Cys32, 35, 62, 65, 73) availability. Moreover, Hg-thiol interaction also is crucial in the neurotoxicity of Hg by modulating the cytoskeleton and neuronal receptors, to name a few. However, existing data on the role of Hg-SH binding in the Hg toxicity remains poorly defined. Therefore, more research is needed to understand better the role of Hg-thiol binding in the molecular pathways of Hg toxicology and the critical role of thiols to counteract negative effects of Hg overload.
Keywords: Apoptosis; Conjugates; Cysteine; Mercury; S-mercuration.
Conflict of interest statement
Declaration of Competing Interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
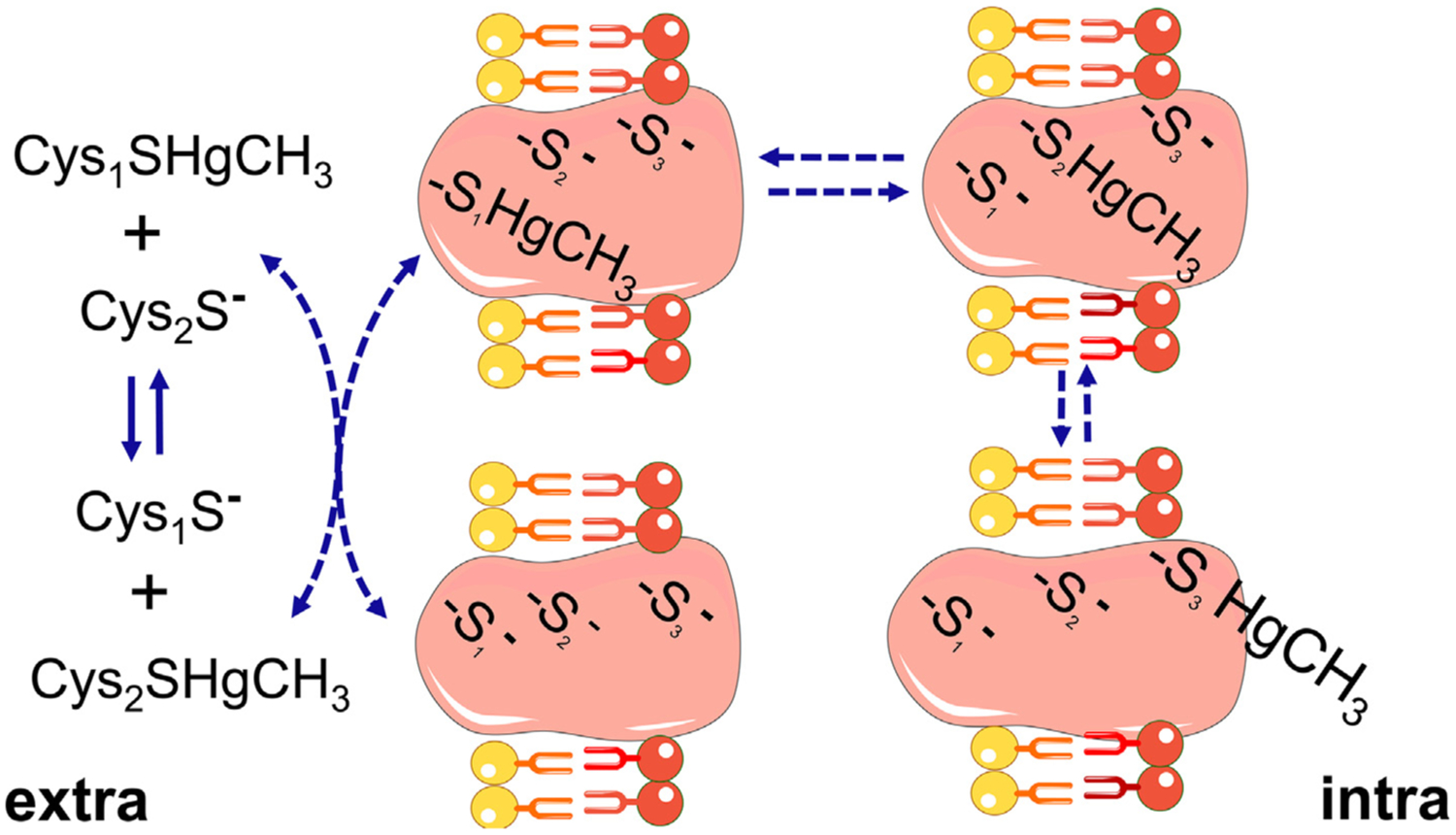
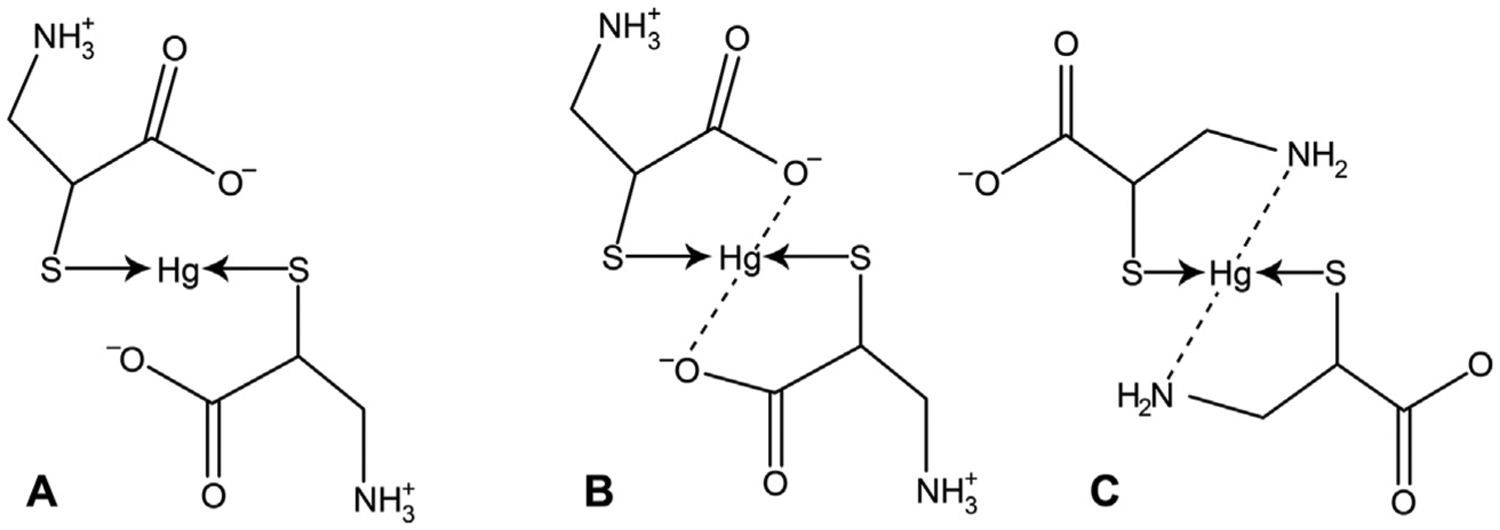
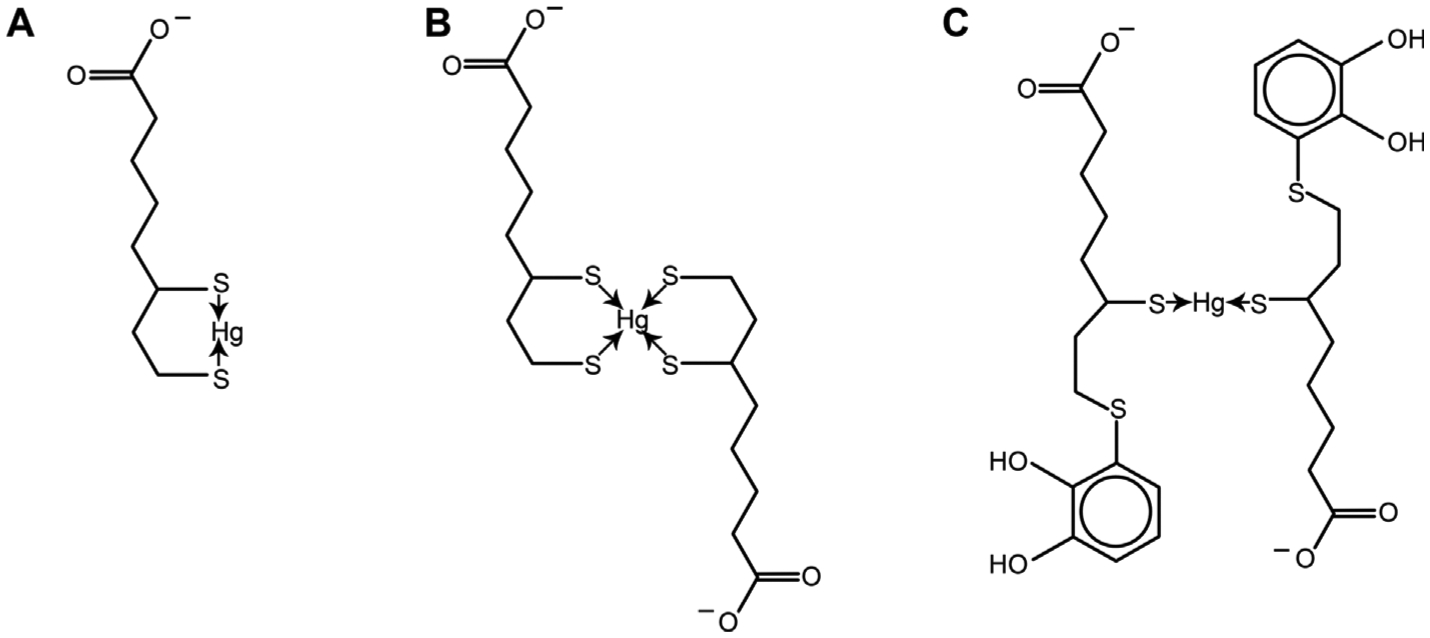
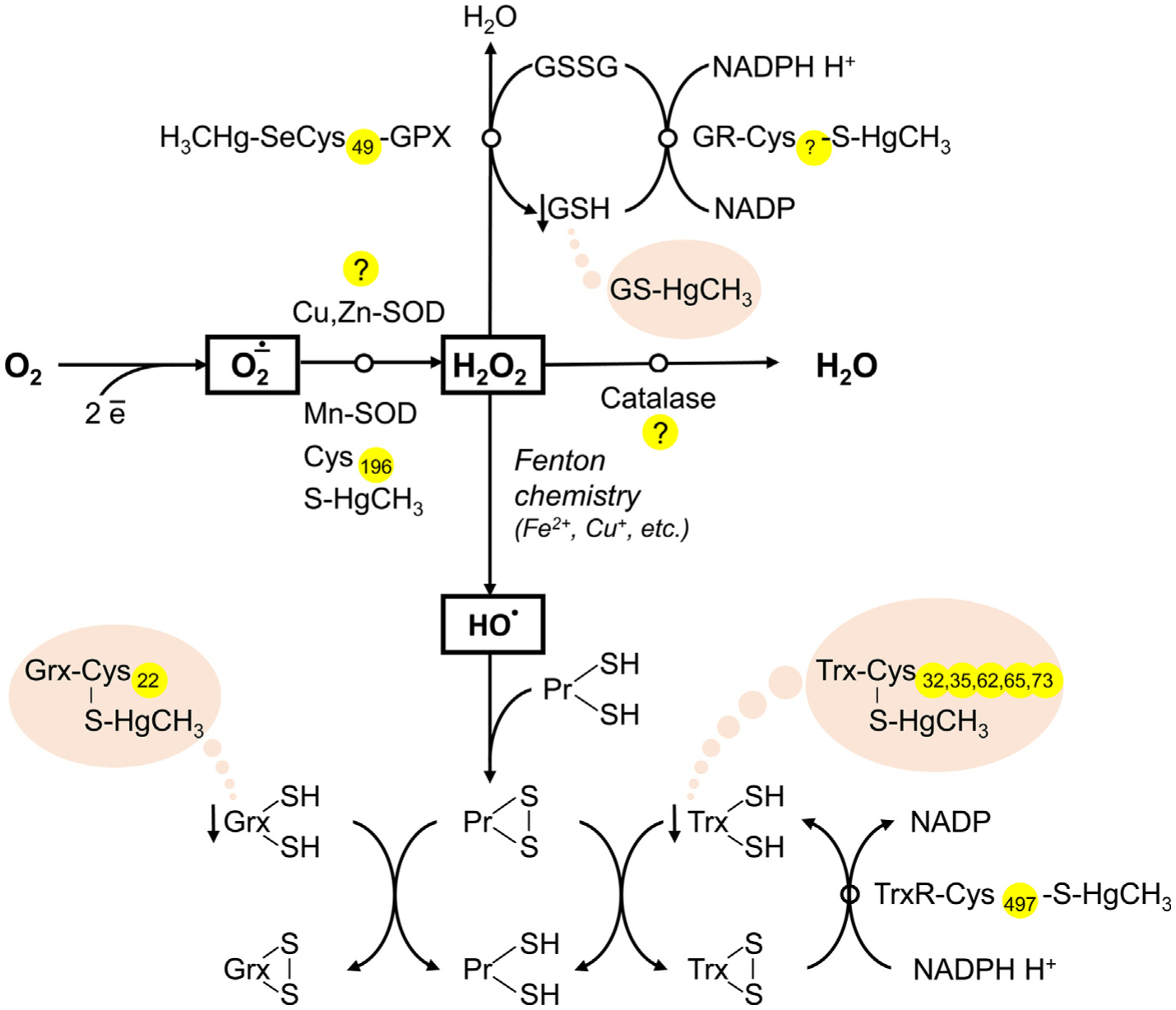
References
-
- International Programme on Chemical Safety, in, World Health Organization, Geneva, 2010.
-
- Evers DC, Keane SE, Basu N, Buck D, Sci. Total Environ 569 (2016) 888–903. - PubMed
-
- Streets DG, Horowitz HM, Lu Z, Levin L, Thackray CP, Sunderland EM, Atmos. Environ 201 (2019) 417–427.
-
- Pacyna EG, Pacyna J, Sundseth K, Munthe J, Kindbom K, Wilson S, Steenhuisen F, Maxson P, Atmos. Environ 44 (2010) 2487–2499.
Grants and funding
LinkOut - more resources
Full Text Sources