

multicrispr: gRNA design for prime editing and parallel targeting of thousands of targets
- PMID: 32907859
- PMCID: PMC7494814
- DOI: 10.26508/lsa.202000757
multicrispr: gRNA design for prime editing and parallel targeting of thousands of targets
Abstract
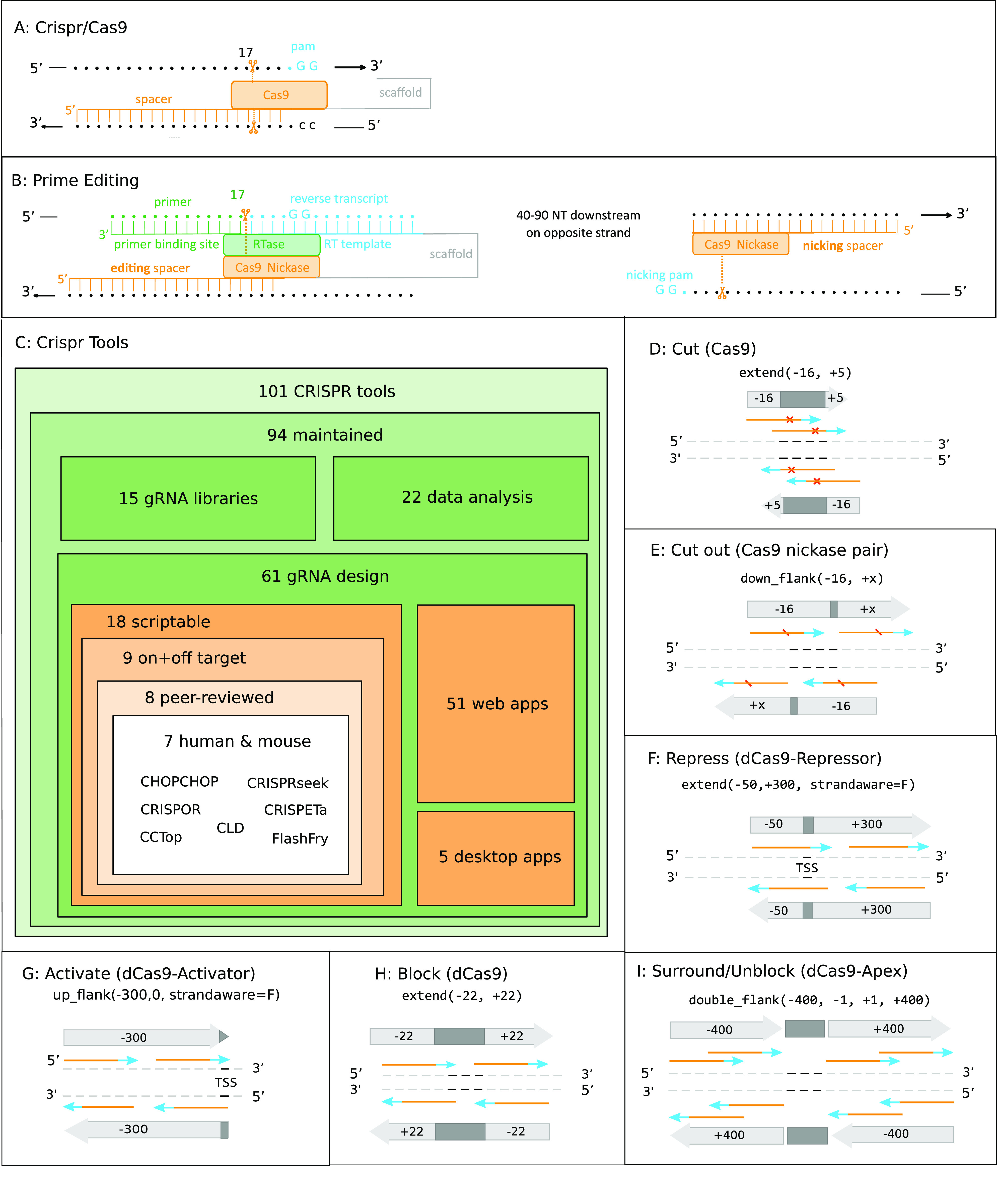
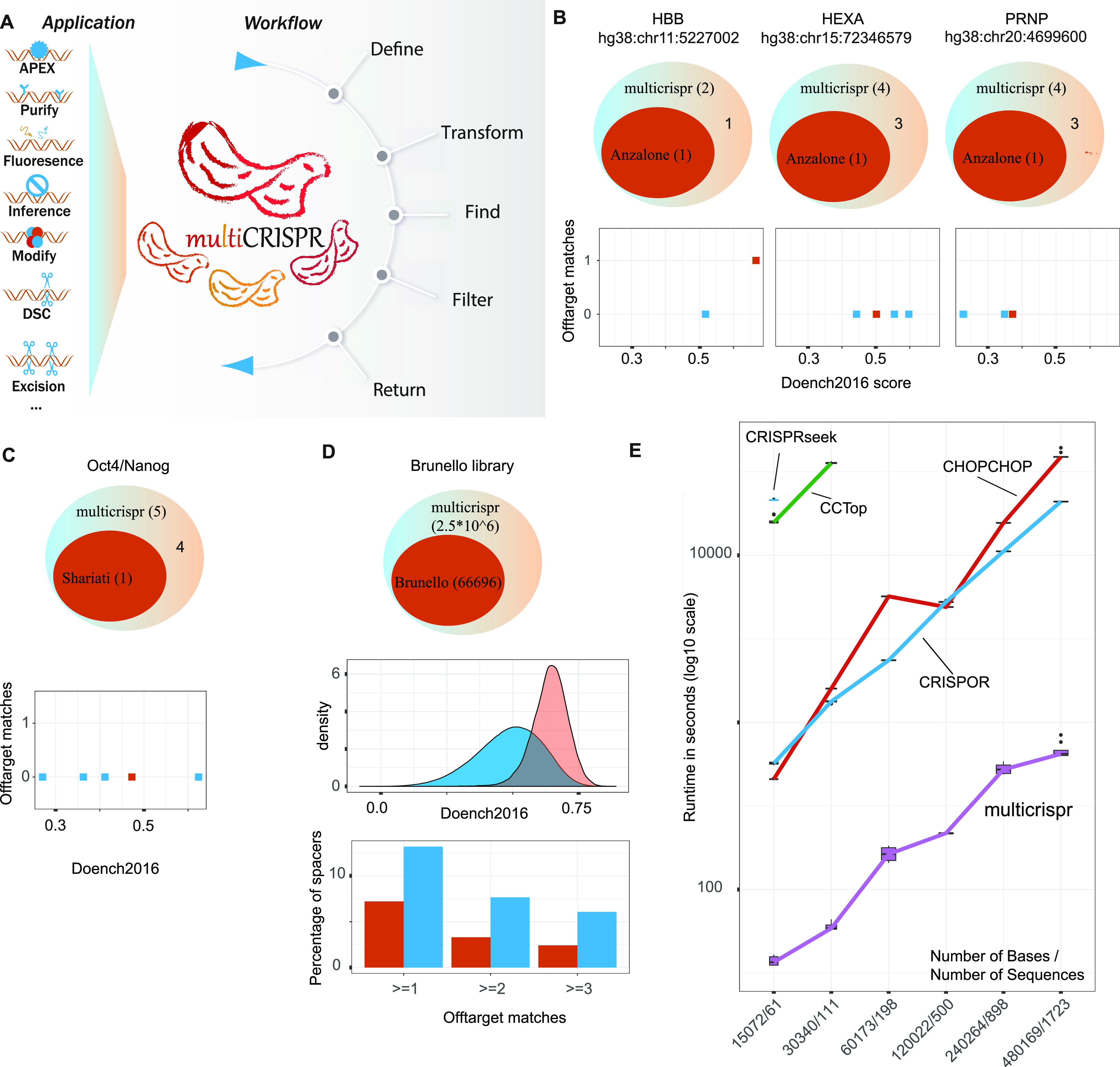
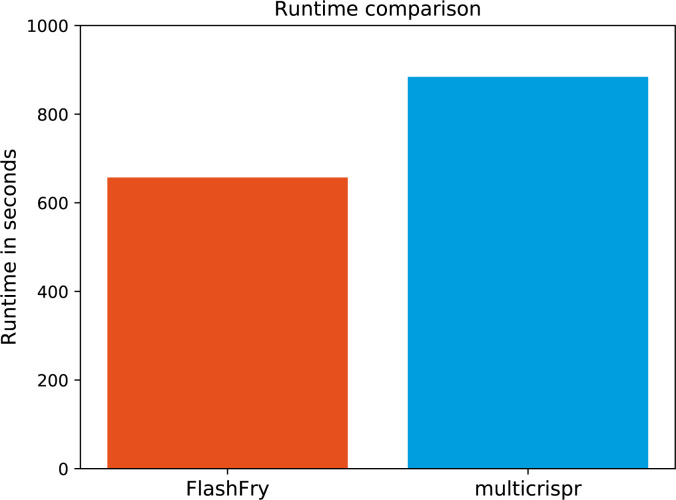
Targeting the coding genome to introduce nucleotide deletions/insertions via the CRISPR/Cas9 technology has become a standard procedure. It has quickly spawned a multitude of methods such as prime editing, APEX proximity labeling, or homology directed repair, for which supporting bioinformatics tools are, however, lagging behind. New CRISPR/Cas9 applications often require specific gRNA design functionality, and a generic tool is critically missing. Here, we introduce multicrispr, an R/bioconductor tool, intended to design individual gRNAs and complex gRNA libraries. The package is easy to use; detects, scores, and filters gRNAs on both efficiency and specificity; visualizes and aggregates results per target or CRISPR/Cas9 sequence; and finally returns both genomic ranges and sequences of gRNAs. To be generic, multicrispr defines and implements a genomic arithmetic framework as a basis for facile adaptation to techniques recently introduced such as prime editing or yet to arise. Its performance and design concepts such as target set-specific filtering render multicrispr a tool of choice when dealing with screening-like approaches.
© 2020 Bhagwat et al.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

References
-
- Aho AV, Corasick MJ (1975) Efficient string matching: An aid to bibliographic search. Commun ACM 18: 333–340. 10.1145/360825.360855 MR 0371172 - DOI
-
- Anaconda Development Team (2016) Anaconda Software Distribution. Computer software. Version 2-2.4.0. Anaconda.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous