

Allosteric regulation of lysosomal enzyme recognition by the cation-independent mannose 6-phosphate receptor
- PMID: 32908216
- PMCID: PMC7481795
- DOI: 10.1038/s42003-020-01211-w
Allosteric regulation of lysosomal enzyme recognition by the cation-independent mannose 6-phosphate receptor
Abstract
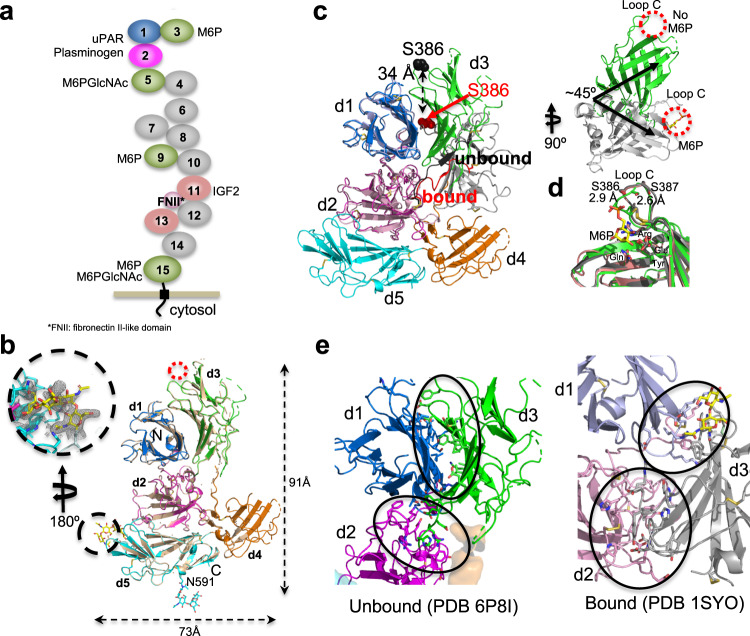
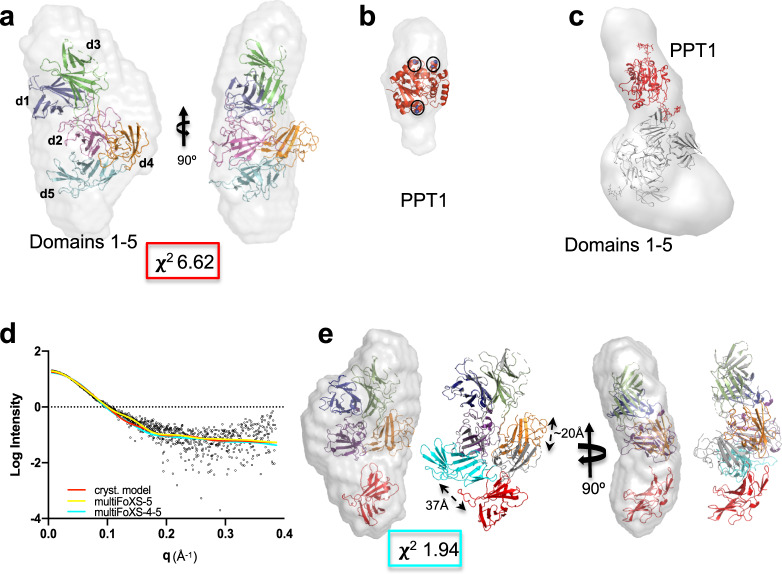
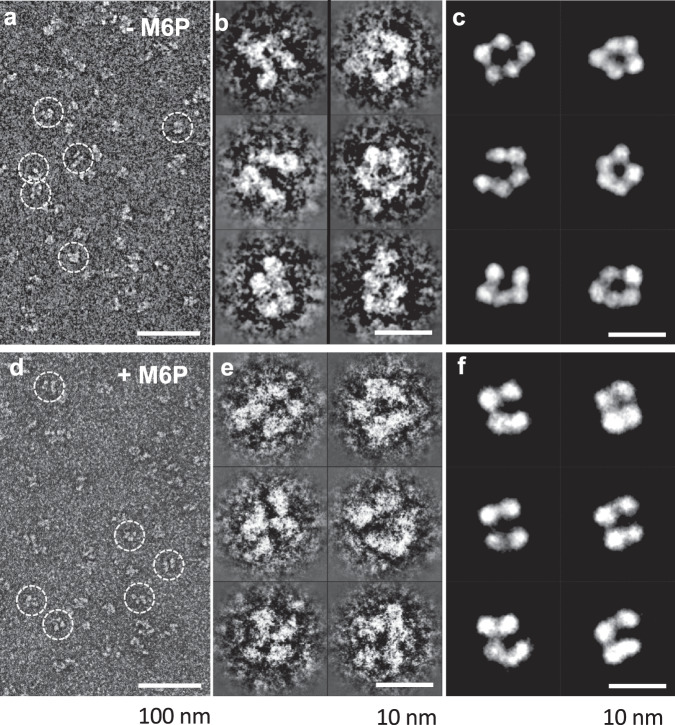
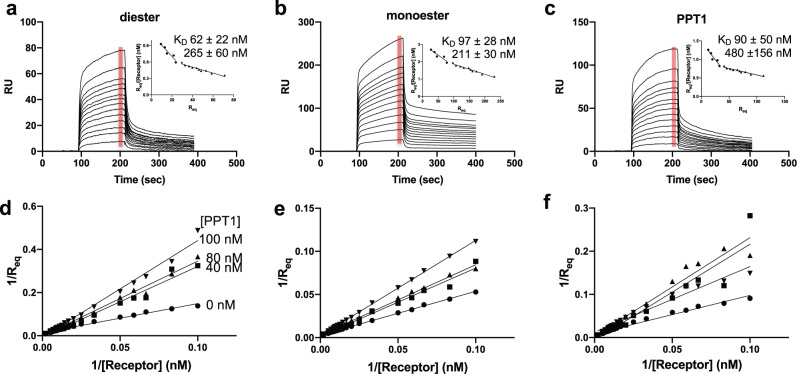
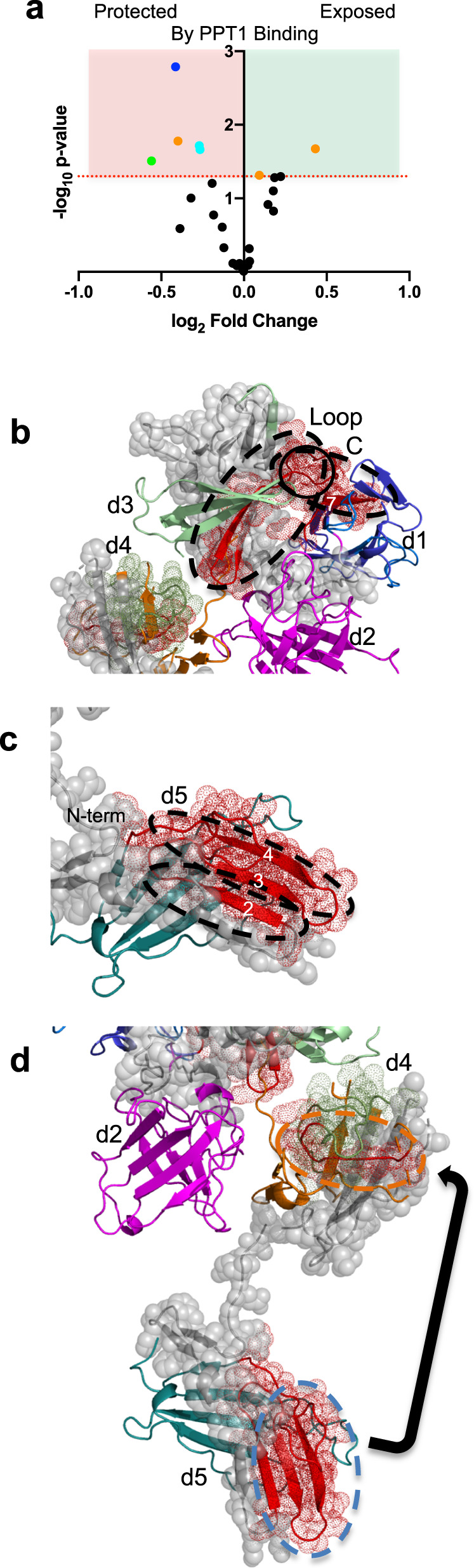
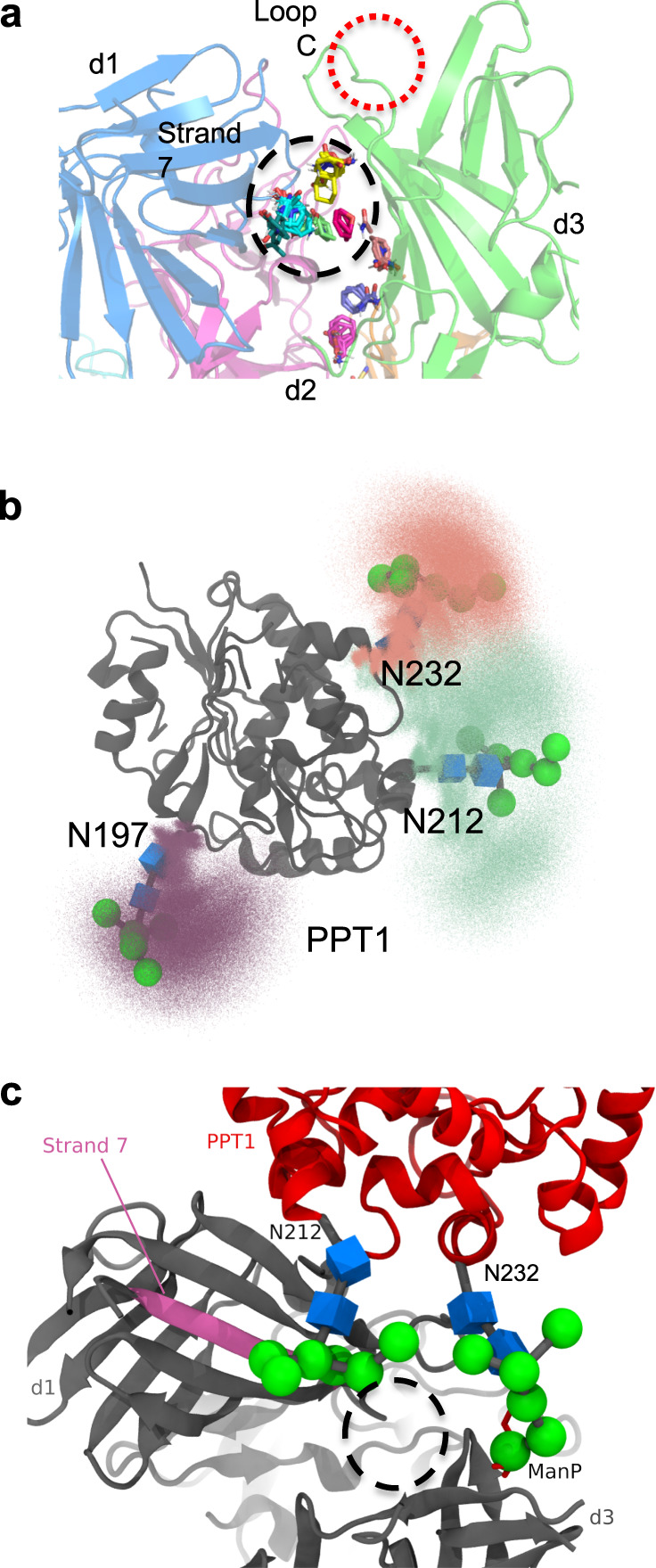
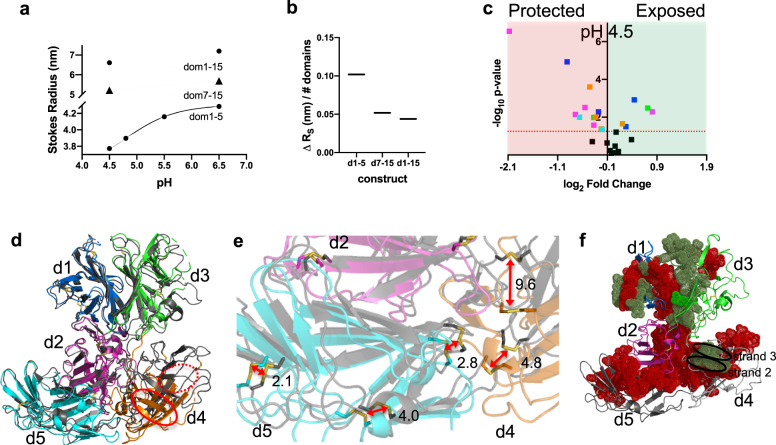
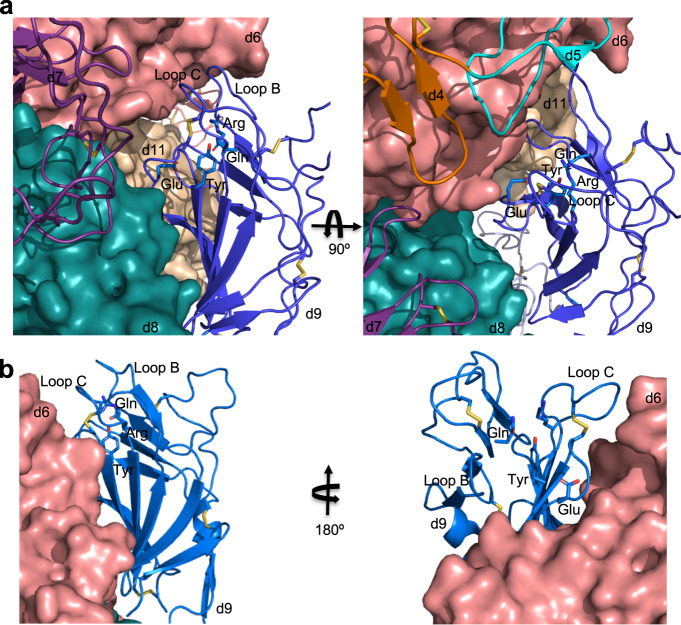
The cation-independent mannose 6-phosphate receptor (CI-MPR, IGF2 receptor or CD222), is a multifunctional glycoprotein required for normal development. Through the receptor's ability to bind unrelated extracellular and intracellular ligands, it participates in numerous functions including protein trafficking, lysosomal biogenesis, and regulation of cell growth. Clinically, endogenous CI-MPR delivers infused recombinant enzymes to lysosomes in the treatment of lysosomal storage diseases. Although four of the 15 domains comprising CI-MPR's extracellular region bind phosphorylated glycans on lysosomal enzymes, knowledge of how CI-MPR interacts with ~60 different lysosomal enzymes is limited. Here, we show by electron microscopy and hydroxyl radical protein footprinting that the N-terminal region of CI-MPR undergoes dynamic conformational changes as a consequence of ligand binding and different pH conditions. These data, coupled with X-ray crystallography, surface plasmon resonance and molecular modeling, allow us to propose a model explaining how high-affinity carbohydrate binding is achieved through allosteric domain cooperativity.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Ghosh P, Dahms NM, Kornfeld S. Mannose 6-phosphate receptors: new twists in the tale. Nat. Rev. Mol. Cell Biol. 2003;4:202–212. - PubMed
-
- El-Shewy HM, Luttrell LM. Insulin-like growth factor-2/mannose-6 phosphate receptors. Vitam. Horm. 2009;80:667–697. - PubMed
-
- Blanchard F, et al. The mannose 6-phosphate/insulin-like growth factor II receptor is a nanomolar affinity receptor for glycosylated human leukemia inhibitory factor. J. Biol. Chem. 1998;273:20886–20893. - PubMed
-
- Morgan DO, et al. Insulin-like growth factor II receptor as a multifunctional binding protein. Nature. 1987;329:301–307. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
