

Genetic models of human and mouse dendritic cell development and function
- PMID: 32908299
- PMCID: PMC10955724
- DOI: 10.1038/s41577-020-00413-x
Genetic models of human and mouse dendritic cell development and function
Abstract
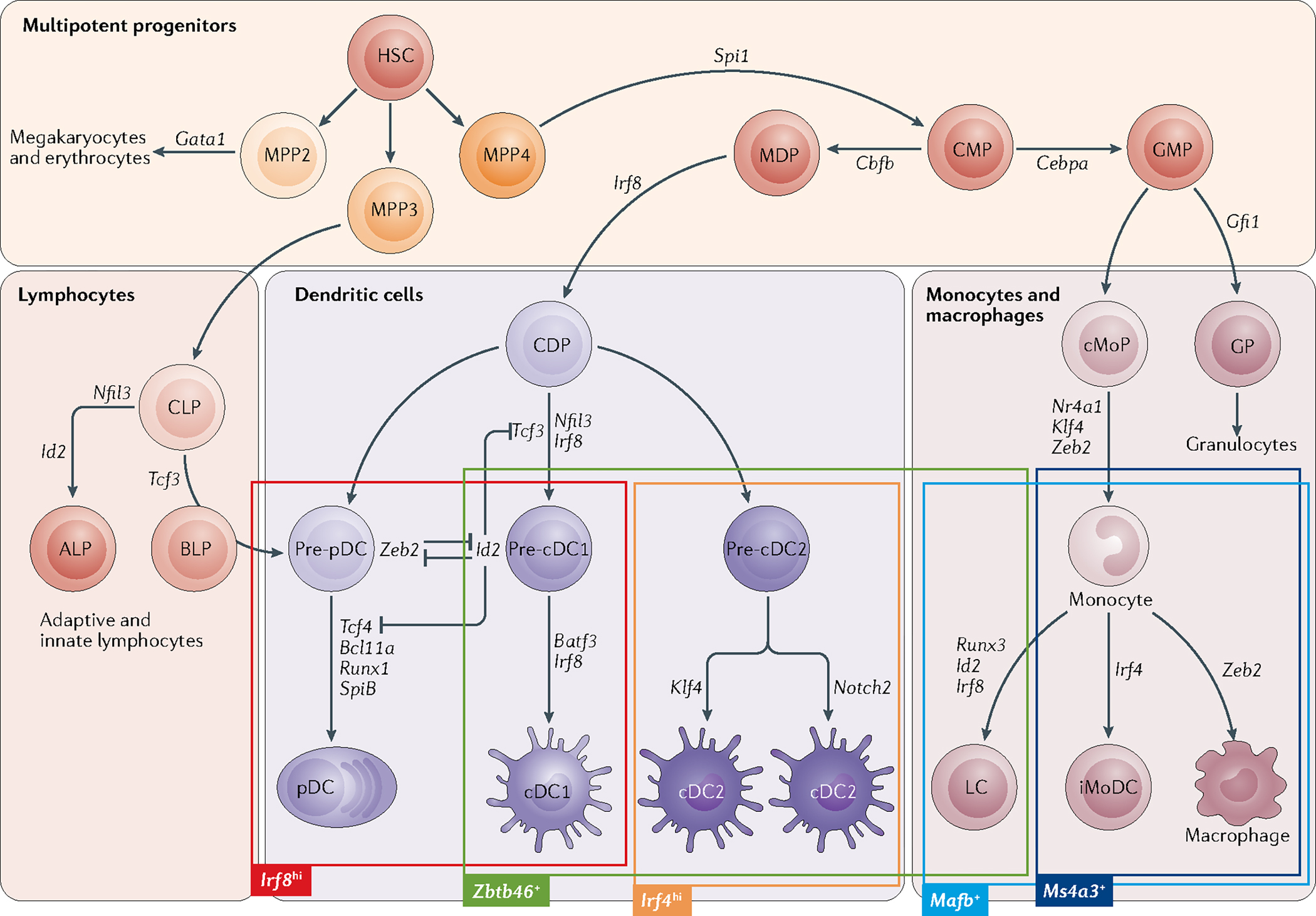
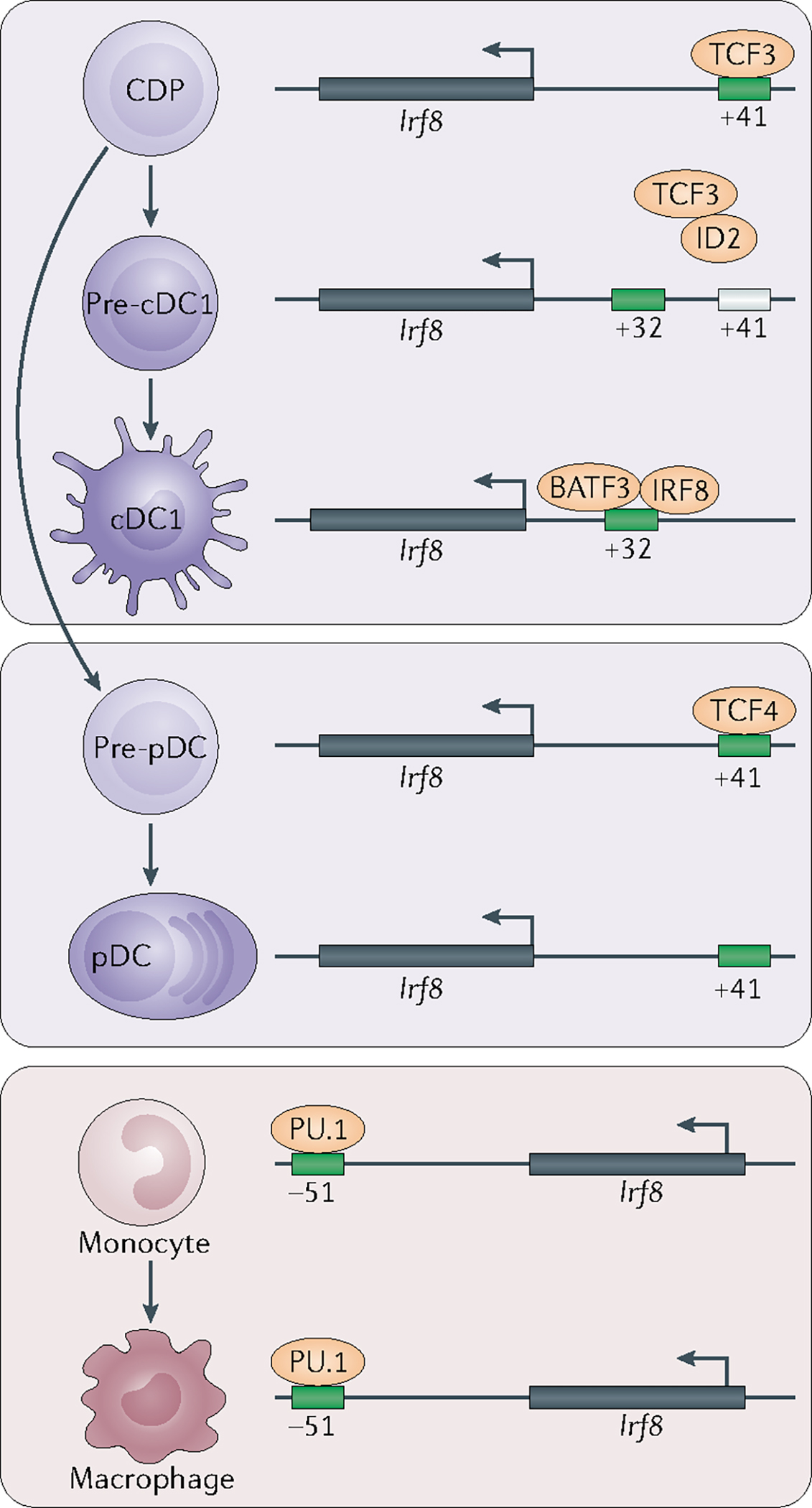
Dendritic cells (DCs) develop in the bone marrow from haematopoietic progenitors that have numerous shared characteristics between mice and humans. Human counterparts of mouse DC progenitors have been identified by their shared transcriptional signatures and developmental potential. New findings continue to revise models of DC ontogeny but it is well accepted that DCs can be divided into two main functional groups. Classical DCs include type 1 and type 2 subsets, which can detect different pathogens, produce specific cytokines and present antigens to polarize mainly naive CD8+ or CD4+ T cells, respectively. By contrast, the function of plasmacytoid DCs is largely innate and restricted to the detection of viral infections and the production of type I interferon. Here, we discuss genetic models of mouse DC development and function that have aided in correlating ontogeny with function, as well as how these findings can be translated to human DCs and their progenitors.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures

References
-
- Dress RJ et al. Plasmacytoid dendritic cells develop from Ly6D+ lymphoid progenitors distinct from the myeloid lineage. Nat. Immunol. 20, 852–864 (2019). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
