

This is a preprint.
Spike mutation D614G alters SARS-CoV-2 fitness and neutralization susceptibility
- PMID: 32908978
- PMCID: PMC7480025
- DOI: 10.1101/2020.09.01.278689
Spike mutation D614G alters SARS-CoV-2 fitness and neutralization susceptibility
Update in
-
Spike mutation D614G alters SARS-CoV-2 fitness.Nature. 2021 Apr;592(7852):116-121. doi: 10.1038/s41586-020-2895-3. Epub 2020 Oct 26. Nature. 2021. PMID: 33106671 Free PMC article.
Abstract
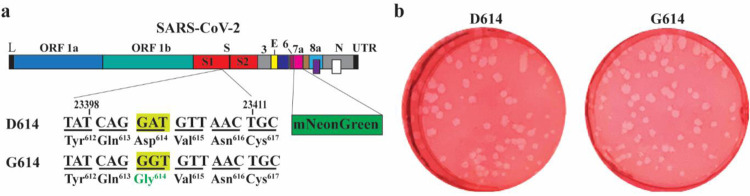
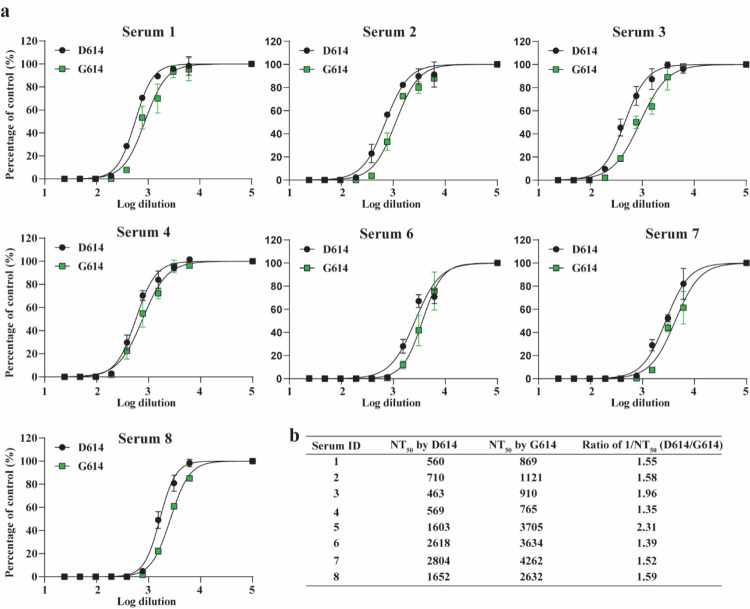
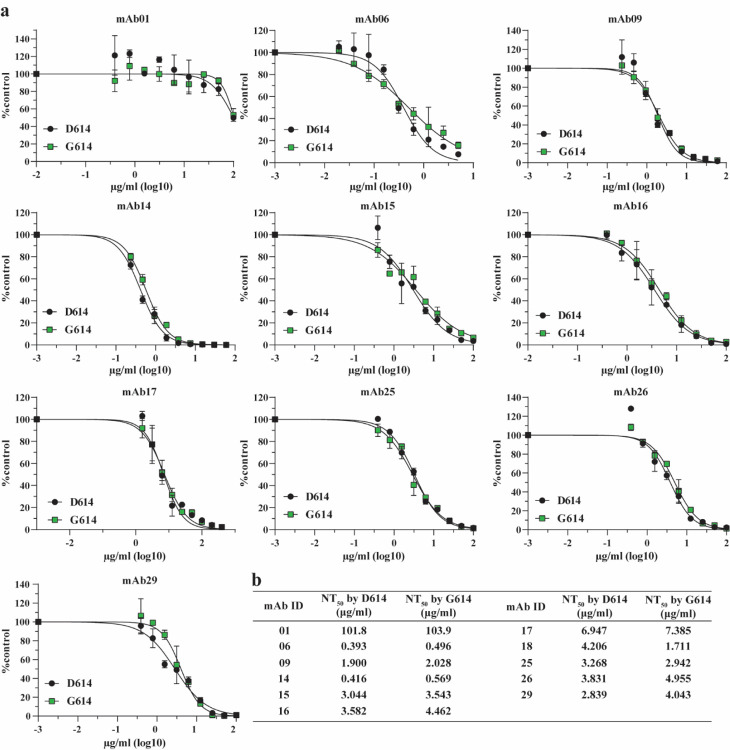
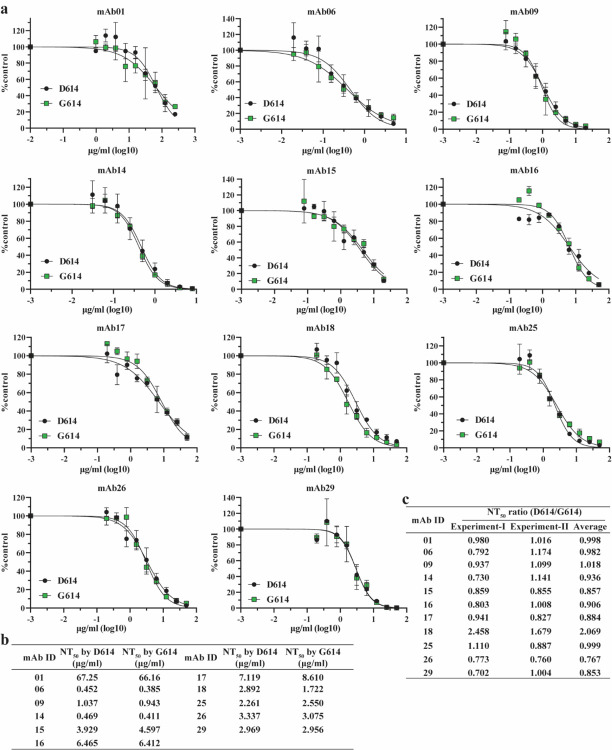
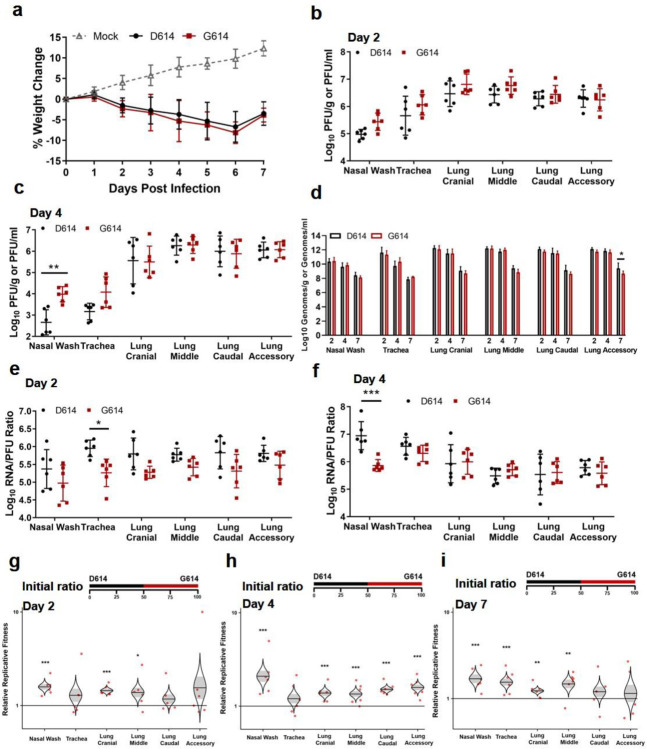
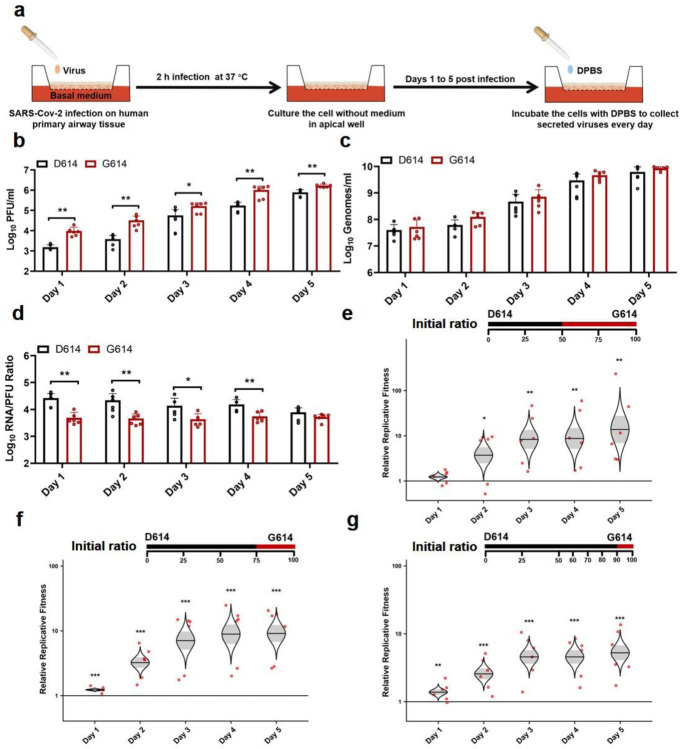
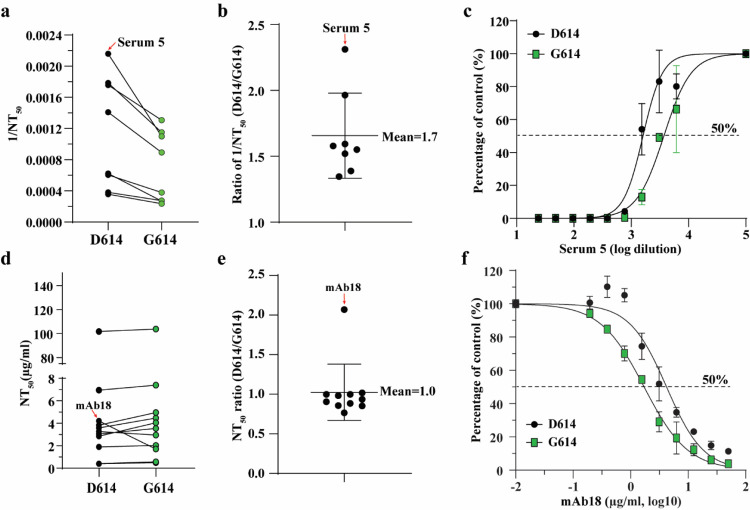
A spike protein mutation D614G became dominant in SARS-CoV-2 during the COVID-19 pandemic. However, the mutational impact on viral spread and vaccine efficacy remains to be defined. Here we engineer the D614G mutation in the SARS-CoV-2 USA-WA1/2020 strain and characterize its effect on viral replication, pathogenesis, and antibody neutralization. The D614G mutation significantly enhances SARS-CoV-2 replication on human lung epithelial cells and primary human airway tissues, through an improved infectivity of virions with the spike receptor-binding domain in an "up" conformation for binding to ACE2 receptor. Hamsters infected with D614 or G614 variants developed similar levels of weight loss. However, the G614 virus produced higher infectious titers in the nasal washes and trachea, but not lungs, than the D614 virus. The hamster results confirm clinical evidence that the D614G mutation enhances viral loads in the upper respiratory tract of COVID-19 patients and may increases transmission. For antibody neutralization, sera from D614 virus-infected hamsters consistently exhibit higher neutralization titers against G614 virus than those against D614 virus, indicating that (i) the mutation may not reduce the ability of vaccines in clinical trials to protect against COVID-19 and (ii) therapeutic antibodies should be tested against the circulating G614 virus before clinical development.
Conflict of interest statement
Competing financial interests X.X., V.D.M., and P.-Y.S. have filed a patent on the reverse genetic system and reporter SARS-CoV-2. Other authors declare no competing interests.
Figures
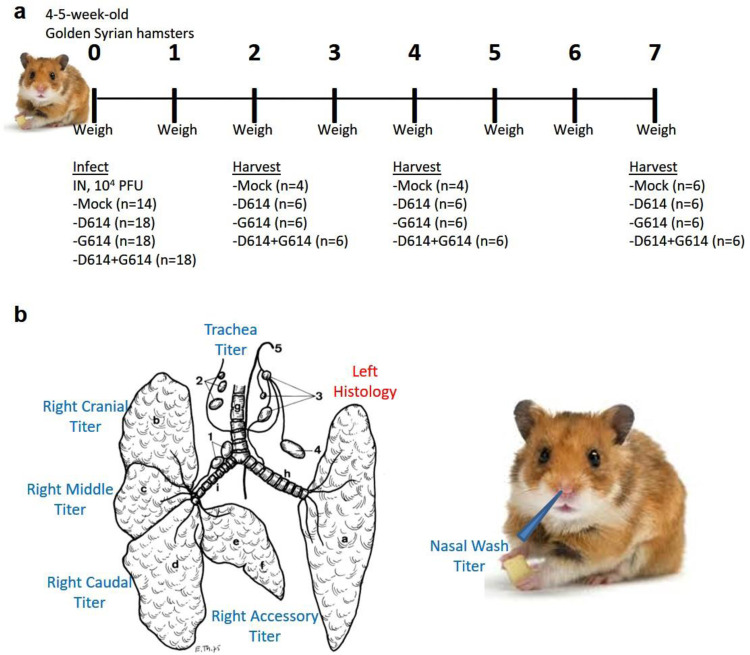
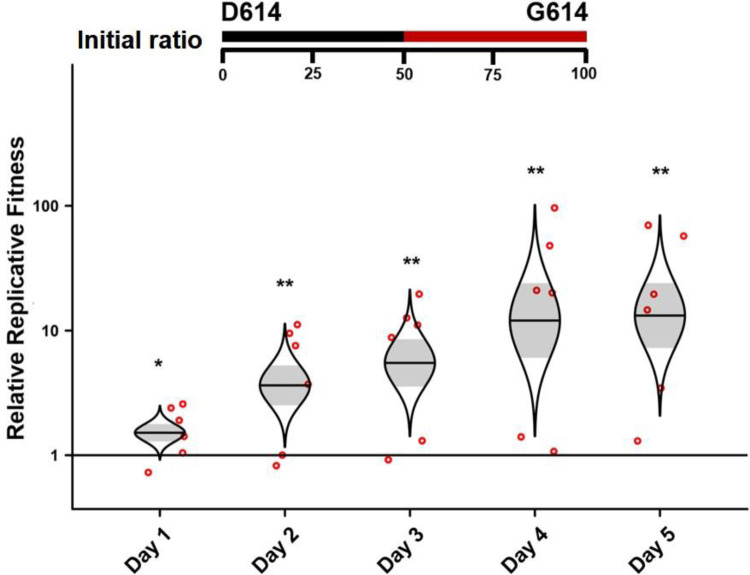
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous