

The Sda Synthase B4GALNT2 Reduces Malignancy and Stemness in Colon Cancer Cell Lines Independently of Sialyl Lewis X Inhibition
- PMID: 32911675
- PMCID: PMC7555213
- DOI: 10.3390/ijms21186558
The Sda Synthase B4GALNT2 Reduces Malignancy and Stemness in Colon Cancer Cell Lines Independently of Sialyl Lewis X Inhibition
Abstract
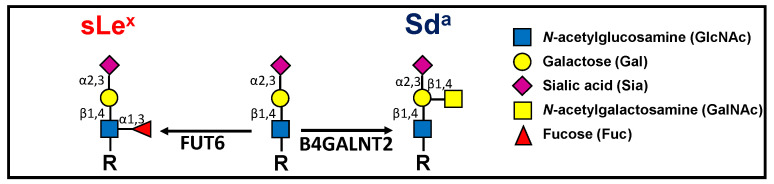
Background: The Sda antigen and its biosynthetic enzyme B4GALNT2 are highly expressed in healthy colon but undergo a variable down-regulation in colon cancer. The biosynthesis of the malignancy-associated sialyl Lewis x (sLex) antigen in normal and cancerous colon is mediated by fucosyltransferase 6 (FUT6) and is mutually exclusive from that of Sda. It is thought that the reduced malignancy associated with high B4GALNT2 was due to sLex inhibition.
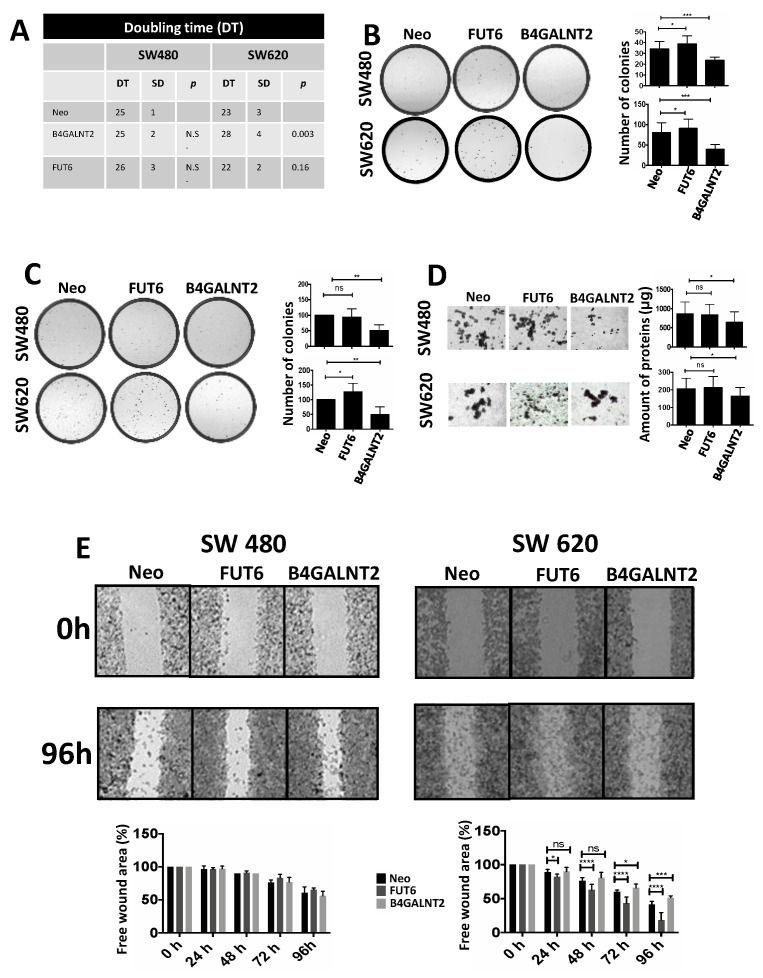
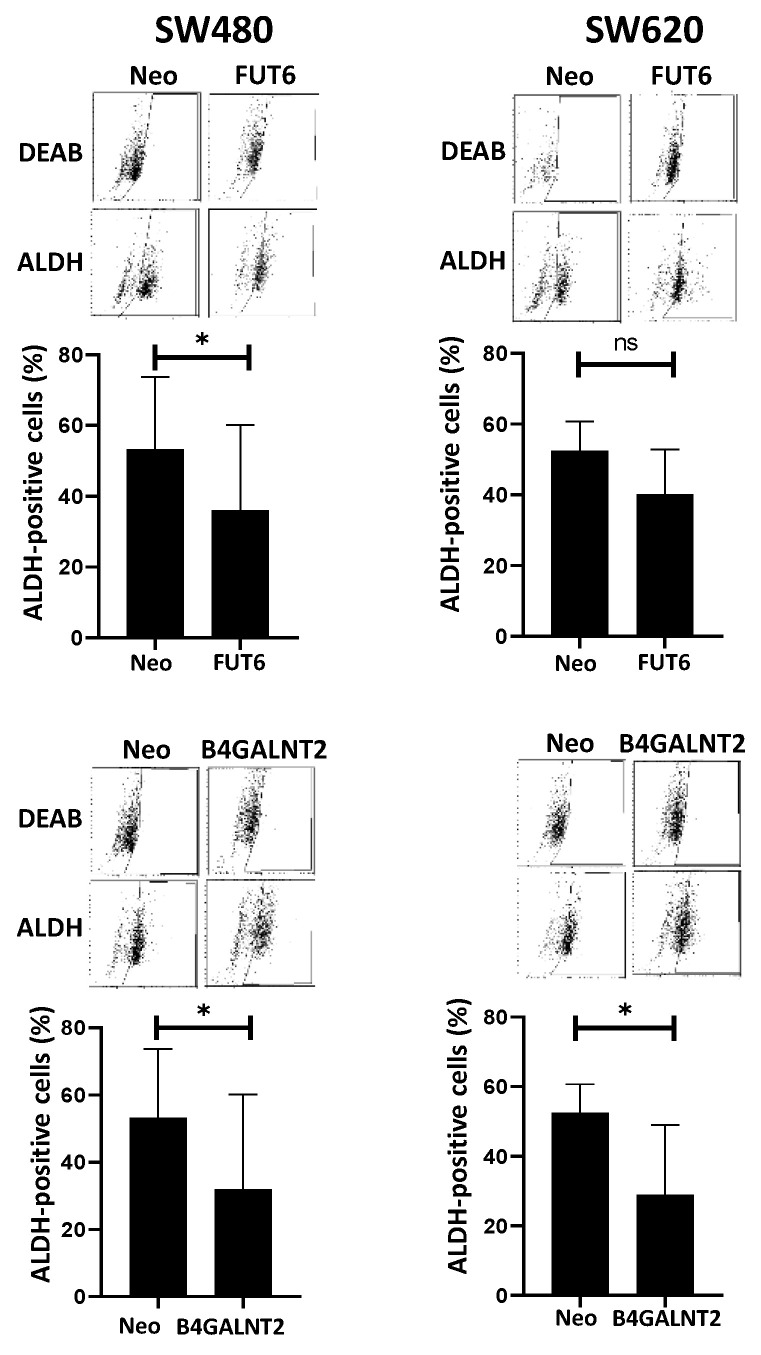
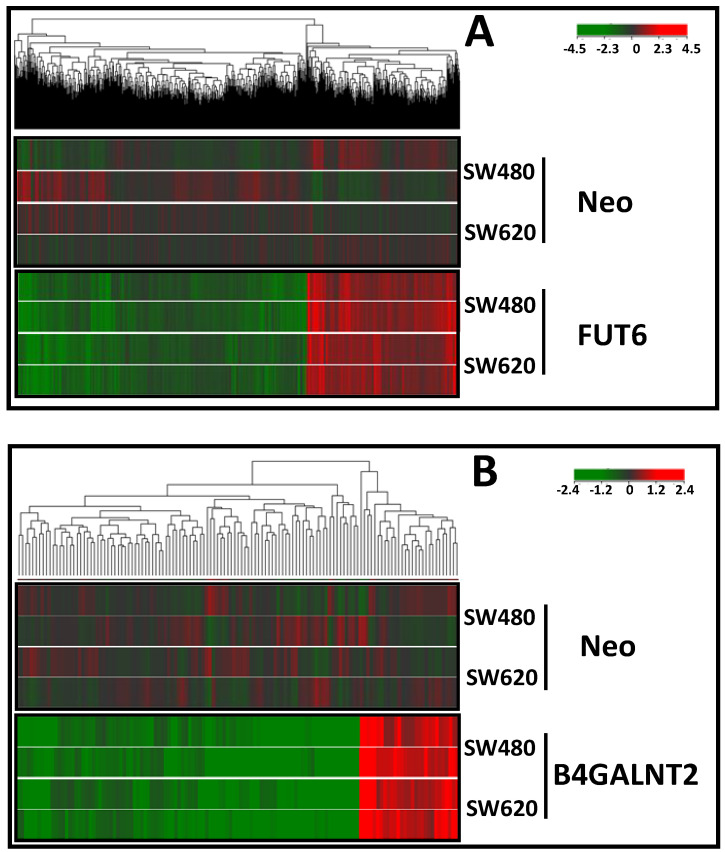
Methods: We transfected the cell lines SW480 and SW620, derived respectively from a primary tumor and a metastasis of the same patient, with the cDNAs of FUT6 or B4GALNT2, generating cell variants expressing either the sLex or the Sda antigens. Transfectants were analyzed for growth in poor adherence, wound healing, stemness and gene expression profile.
Results: B4GALNT2/Sda expression down-regulated all malignancy-associated phenotypes in SW620 but only those associated with stemness in SW480. FUT6/sLex enhanced some malignancy-associated phenotypes in SW620, but had little effect in SW480. The impact on the transcriptome was stronger for FUT6 than for B4GALNT2 and only partially overlapping between SW480 and SW620.
Conclusions: B4GALNT2/Sda inhibits the stemness-associated malignant phenotype, independently of sLex inhibition. The impact of glycosyltransferases on the phenotype and the transcriptome is highly cell-line specific.
Keywords: Sda antigen; cancer stem cells; non-adherent growth; sialyl Lewis antigens; transcriptomic analysis.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
Similar articles
-
The Cancer-Associated Antigens Sialyl Lewisa/x and Sda: Two Opposite Faces of Terminal Glycosylation.Cancers (Basel). 2021 Oct 21;13(21):5273. doi: 10.3390/cancers13215273. Cancers (Basel). 2021. PMID: 34771437 Free PMC article. Review.
-
B4GALNT2 gene expression controls the biosynthesis of Sda and sialyl Lewis X antigens in healthy and cancer human gastrointestinal tract.Int J Biochem Cell Biol. 2014 Aug;53:442-9. doi: 10.1016/j.biocel.2014.06.009. Epub 2014 Jun 19. Int J Biochem Cell Biol. 2014. PMID: 24953560
-
B4GALNT2 Controls Sda and SLex Antigen Biosynthesis in Healthy and Cancer Human Colon.Chembiochem. 2021 Dec 10;22(24):3381-3390. doi: 10.1002/cbic.202100363. Epub 2021 Sep 14. Chembiochem. 2021. PMID: 34397142 Free PMC article. Review.
-
High Expression of the Sda Synthase B4GALNT2 Associates with Good Prognosis and Attenuates Stemness in Colon Cancer.Cells. 2020 Apr 11;9(4):948. doi: 10.3390/cells9040948. Cells. 2020. PMID: 32290493 Free PMC article.
-
The story of the Sda antigen and of its cognate enzyme B4GALNT2: What is new?Glycoconj J. 2023 Feb;40(1):123-133. doi: 10.1007/s10719-022-10089-1. Epub 2022 Oct 26. Glycoconj J. 2023. PMID: 36287346 Review.
Cited by
-
An Identification of Functional Genetic Variants in B4GALNT2 Gene and Their Association with Growth Traits in Goats.Genes (Basel). 2024 Mar 3;15(3):330. doi: 10.3390/genes15030330. Genes (Basel). 2024. PMID: 38540389 Free PMC article.
-
AC010973.2 promotes cell proliferation and is one of six stemness-related genes that predict overall survival of renal clear cell carcinoma.Sci Rep. 2022 Mar 11;12(1):4272. doi: 10.1038/s41598-022-07070-1. Sci Rep. 2022. PMID: 35277527 Free PMC article.
-
The Cancer-Associated Antigens Sialyl Lewisa/x and Sda: Two Opposite Faces of Terminal Glycosylation.Cancers (Basel). 2021 Oct 21;13(21):5273. doi: 10.3390/cancers13215273. Cancers (Basel). 2021. PMID: 34771437 Free PMC article. Review.
-
N-Glycan on the Non-Consensus N-X-C Glycosylation Site Impacts Activity, Stability, and Localization of the Sda Synthase B4GALNT2.Int J Mol Sci. 2023 Feb 18;24(4):4139. doi: 10.3390/ijms24044139. Int J Mol Sci. 2023. PMID: 36835549 Free PMC article.
-
B4GALNT2 Gene Promotes Proliferation, and Invasiveness and Migration Abilities of Model Triple Negative Breast Cancer (TNBC) Cells by Interacting With HLA-B Protein.Front Oncol. 2021 Sep 13;11:722828. doi: 10.3389/fonc.2021.722828. eCollection 2021. Front Oncol. 2021. PMID: 34589428 Free PMC article.
References
-
- Dall’Olio F., Malagolini N., Chiricolo M. Glycosylation in cancer. Spec. Period. Rep. Carbohydr. Chem. 2012;37:21–56.
-
- Nakamori S., Kameyama M., Imaoka S., Furukawa H., Ishikawa O., Sasaki Y., Kabuto T., Iwanaga T., Matsushita Y., Irimura T. Increased expression of sialyl Lewisx antigen correlates with poor survival in patients with colorectal carcinoma: Clinicopathological and immunohistochemical study. Cancer Res. 1993;53:3632–3637. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
