

Monitoring of XRN4 Targets Reveals the Importance of Cotranslational Decay during Arabidopsis Development
- PMID: 32913043
- PMCID: PMC7608176
- DOI: 10.1104/pp.20.00942
Monitoring of XRN4 Targets Reveals the Importance of Cotranslational Decay during Arabidopsis Development
Abstract
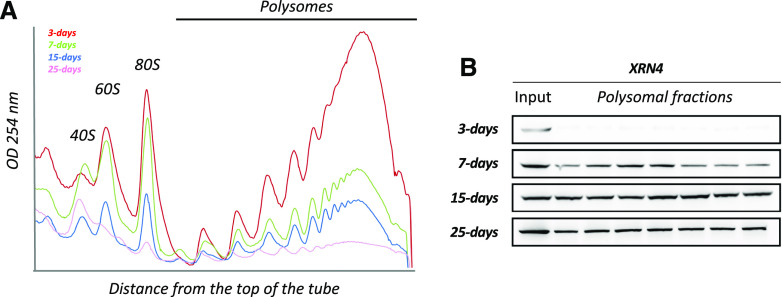
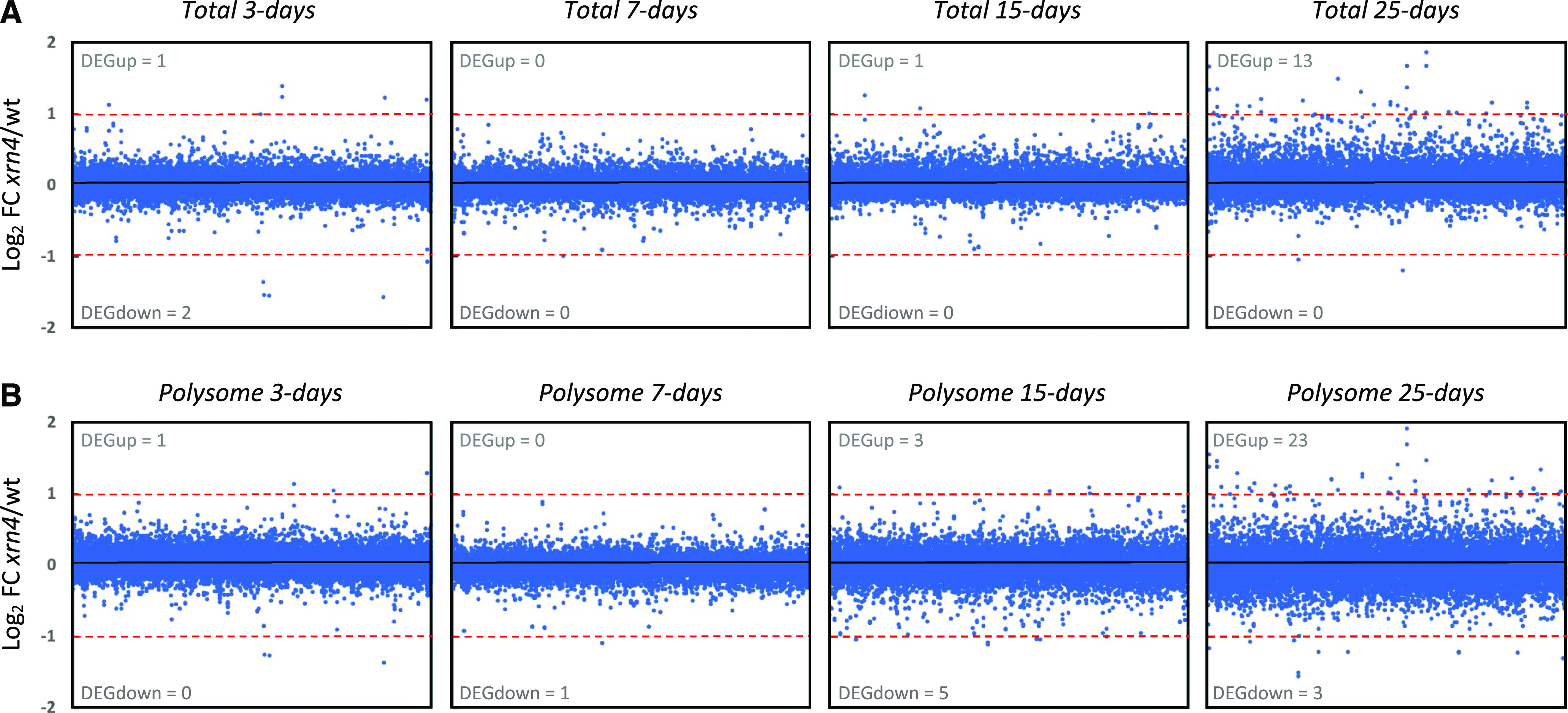
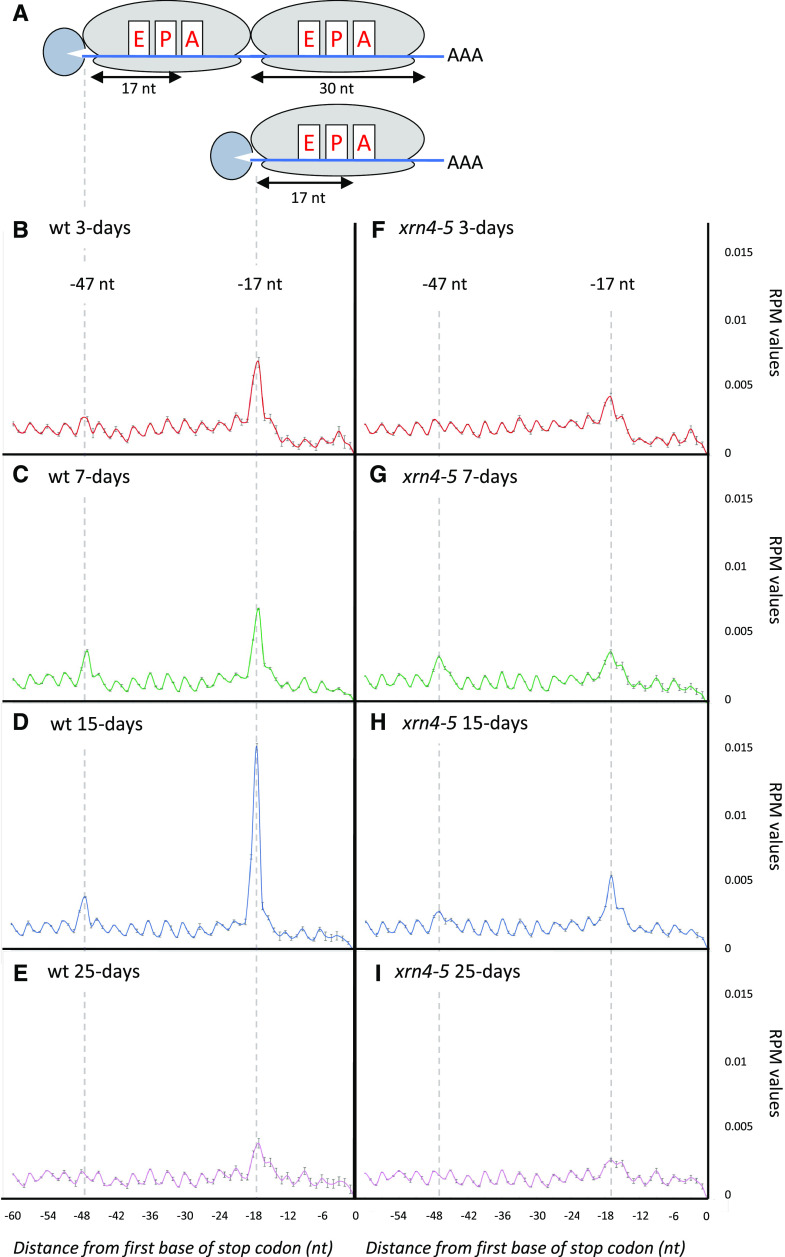
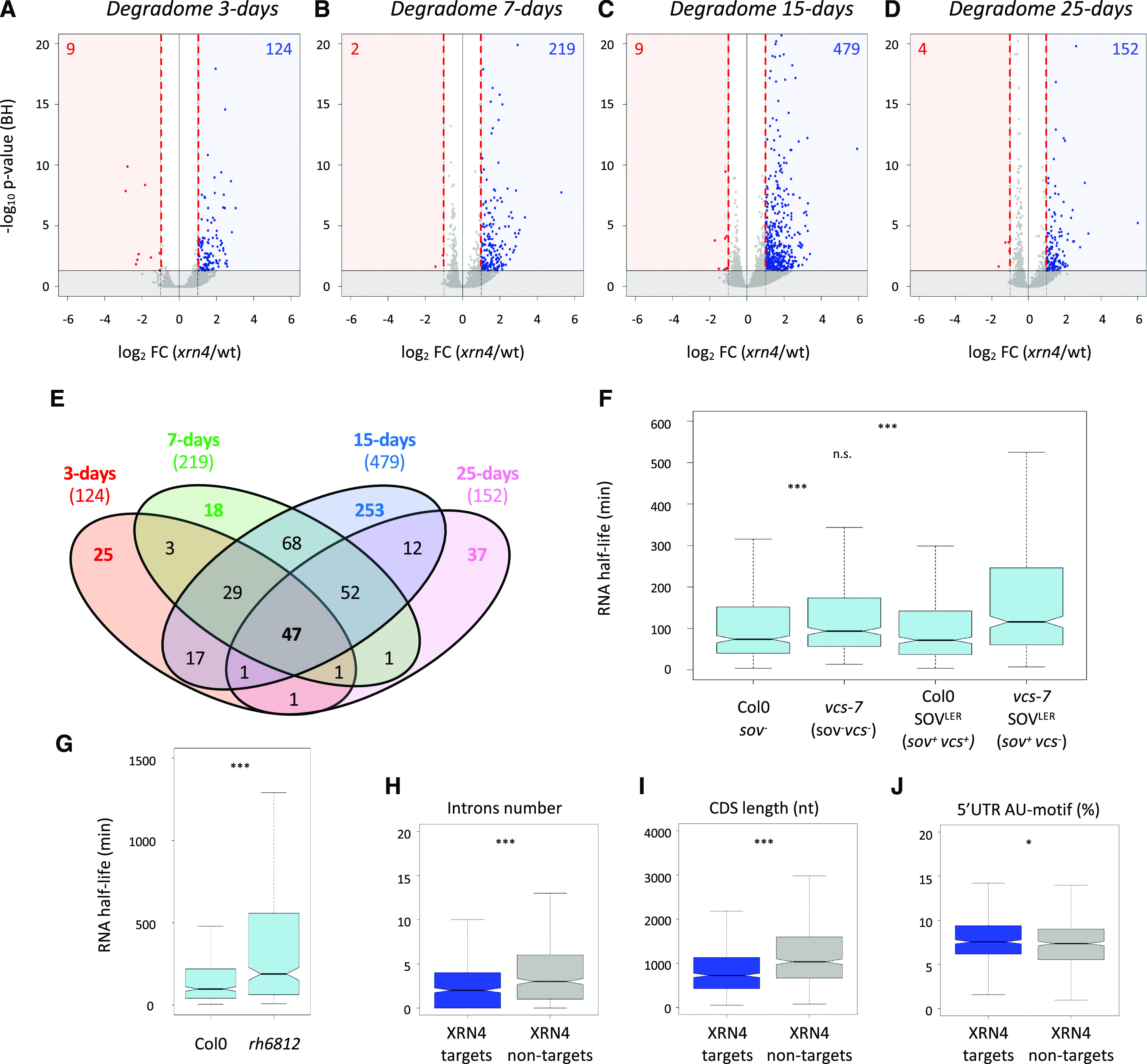
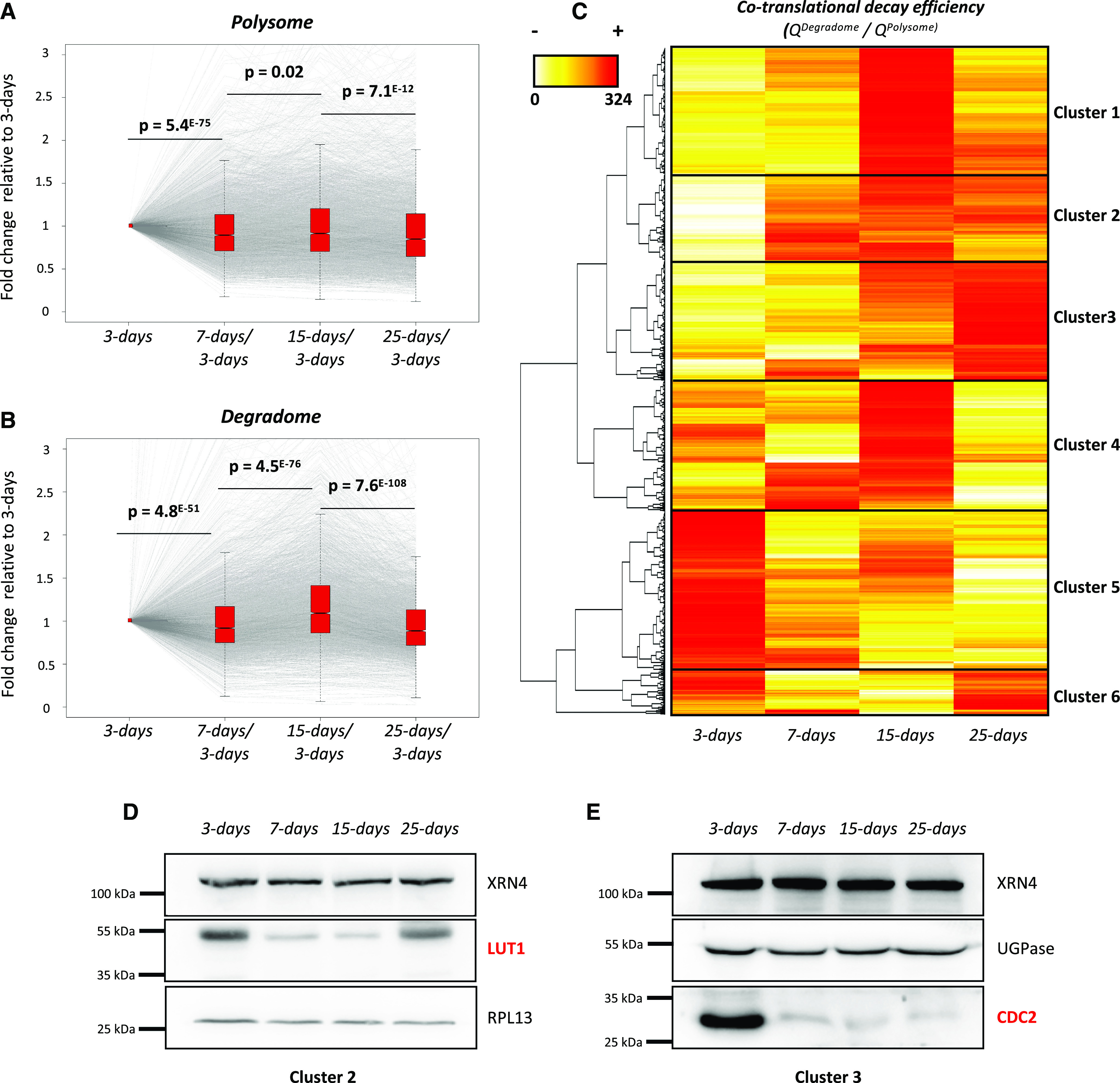
RNA turnover is a general process that maintains appropriate mRNA abundance at the posttranscriptional level. Although long thought to be antagonistic to translation, discovery of the 5' to 3' cotranslational mRNA decay pathway demonstrated that both processes are intertwined. Cotranslational mRNA decay globally shapes the transcriptome in different organisms and in response to stress; however, the dynamics of this process during plant development is poorly understood. In this study, we used a multiomics approach to reveal the global landscape of cotranslational mRNA decay during Arabidopsis (Arabidopsis thaliana) seedling development. We demonstrated that cotranslational mRNA decay is regulated by developmental cues. Using the EXORIBONUCLEASE4 (XRN4) loss-of-function mutant, we showed that XRN4 poly(A+) mRNA targets are largely subject to cotranslational decay during plant development. As cotranslational mRNA decay is interconnected with translation, we also assessed its role in translation efficiency. We discovered that clusters of transcripts were specifically subjected to cotranslational decay in a developmental-dependent manner to modulate their translation efficiency. Our approach allowed the determination of a cotranslational decay efficiency that could be an alternative to other methods to assess transcript translation efficiency. Thus, our results demonstrate the prevalence of cotranslational mRNA decay in plant development and its role in translational control.
© 2020 American Society of Plant Biologists. All Rights Reserved.
Figures
Similar articles
-
Genome-Wide Mapping of Uncapped and Cleaved Transcripts Reveals a Role for the Nuclear mRNA Cap-Binding Complex in Cotranslational RNA Decay in Arabidopsis.Plant Cell. 2016 Oct;28(10):2385-2397. doi: 10.1105/tpc.16.00456. Epub 2016 Oct 7. Plant Cell. 2016. PMID: 27758893 Free PMC article.
-
Evidence that XRN4, an Arabidopsis homolog of exoribonuclease XRN1, preferentially impacts transcripts with certain sequences or in particular functional categories.RNA. 2011 Mar;17(3):501-11. doi: 10.1261/rna.2467911. Epub 2011 Jan 11. RNA. 2011. PMID: 21224377 Free PMC article.
-
Arabidopsis mRNA decay landscape arises from specialized RNA decay substrates, decapping-mediated feedback, and redundancy.Proc Natl Acad Sci U S A. 2018 Feb 13;115(7):E1485-E1494. doi: 10.1073/pnas.1712312115. Epub 2018 Jan 31. Proc Natl Acad Sci U S A. 2018. PMID: 29386391 Free PMC article.
-
RNA-based regulation in the plant circadian clock.Trends Plant Sci. 2011 Oct;16(10):517-23. doi: 10.1016/j.tplants.2011.06.002. Epub 2011 Jul 23. Trends Plant Sci. 2011. PMID: 21782493 Review.
-
Beyond transcription factors: roles of mRNA decay in regulating gene expression in plants.F1000Res. 2018 Dec 17;7:F1000 Faculty Rev-1940. doi: 10.12688/f1000research.16203.1. eCollection 2018. F1000Res. 2018. PMID: 30613385 Free PMC article. Review.
Cited by
-
RNA interference and turnover in plants -a complex partnership.Front Plant Sci. 2025 Jul 1;16:1608888. doi: 10.3389/fpls.2025.1608888. eCollection 2025. Front Plant Sci. 2025. PMID: 40666303 Free PMC article. Review.
-
RNA degradome analysis reveals DNE1 endoribonuclease is required for the turnover of diverse mRNA substrates in Arabidopsis.Plant Cell. 2023 May 29;35(6):1936-1955. doi: 10.1093/plcell/koad085. Plant Cell. 2023. PMID: 37070465 Free PMC article.
-
Monitoring mRNA Half-Life in Arabidopsis Using Droplet Digital PCR.Plants (Basel). 2022 Oct 5;11(19):2616. doi: 10.3390/plants11192616. Plants (Basel). 2022. PMID: 36235485 Free PMC article.
-
Variations in transfer and ribosomal RNA epitranscriptomic status can adapt eukaryote translation to changing physiological and environmental conditions.RNA Biol. 2021 Oct 15;18(sup1):4-18. doi: 10.1080/15476286.2021.1931756. Epub 2021 Jun 23. RNA Biol. 2021. PMID: 34159889 Free PMC article. Review.
-
Endoribonuclease DNE1 Promotes Ethylene Response by Modulating EBF1/2 mRNA Processing in Arabidopsis.Int J Mol Sci. 2024 Feb 10;25(4):2138. doi: 10.3390/ijms25042138. Int J Mol Sci. 2024. PMID: 38396815 Free PMC article.
References
-
- Anderson SJ, Kramer MC, Gosai SJ, Yu X, Vandivier LE, Nelson ADL, Anderson ZD, Beilstein MA, Fray RG, Lyons E, et al. (2018) N6-Methyladenosine inhibits local ribonucleolytic cleavage to stabilize mRNAs in Arabidopsis. Cell Rep 25: 1146–1157.e3 - PubMed
-
- Chantarachot T, Sorenson RS, Hummel M, Ke H, Kettenburg AT, Chen D, Aiyetiwa K, Dehesh K, Eulgem T, Sieburth LE, et al. (2020) DHH1/DDX6-like RNA helicases maintain ephemeral half-lives of stress-response mRNAs. Nat Plants 6: 675–685 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
