

The GTEx Consortium atlas of genetic regulatory effects across human tissues
- PMID: 32913098
- PMCID: PMC7737656
- DOI: 10.1126/science.aaz1776
The GTEx Consortium atlas of genetic regulatory effects across human tissues
Abstract
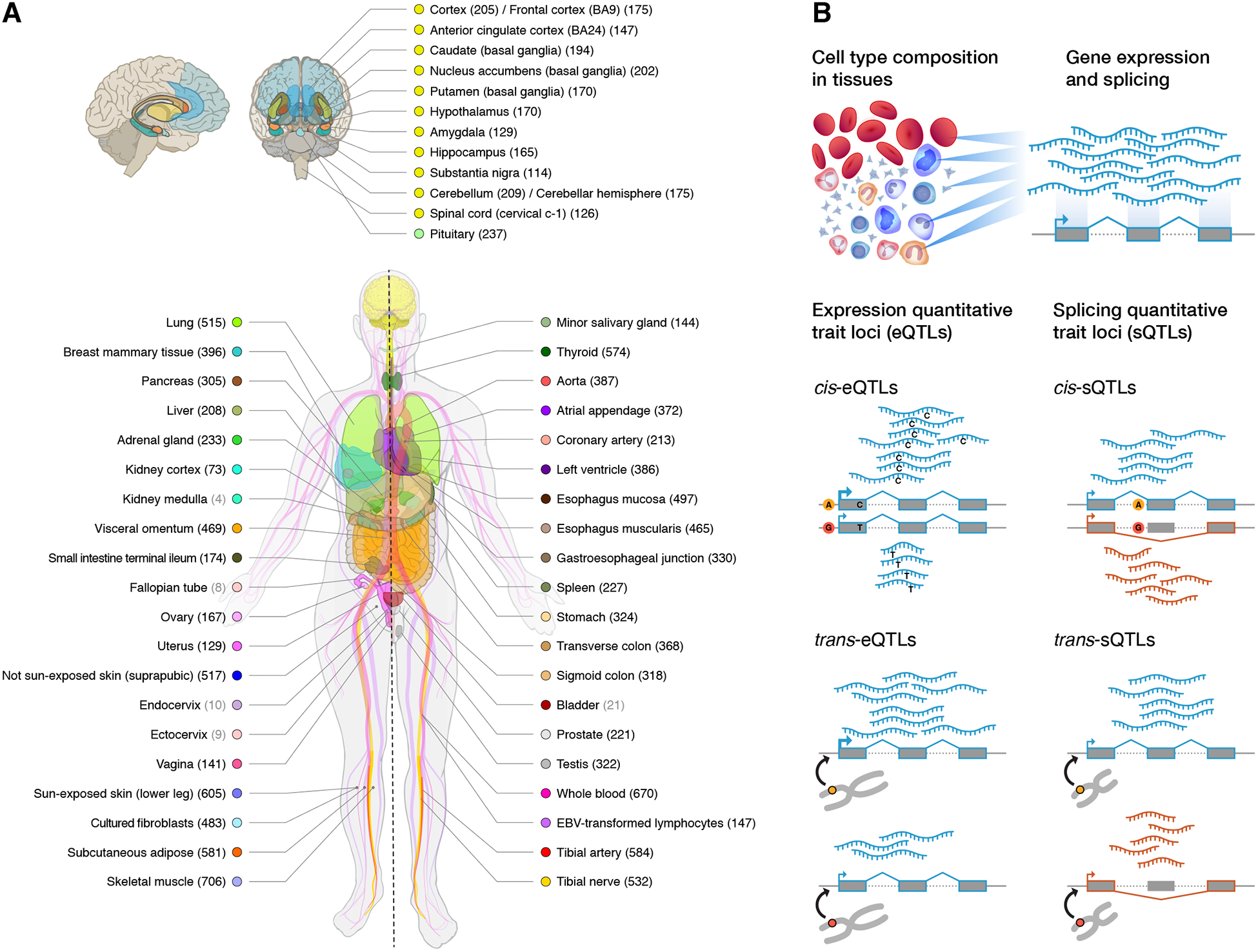
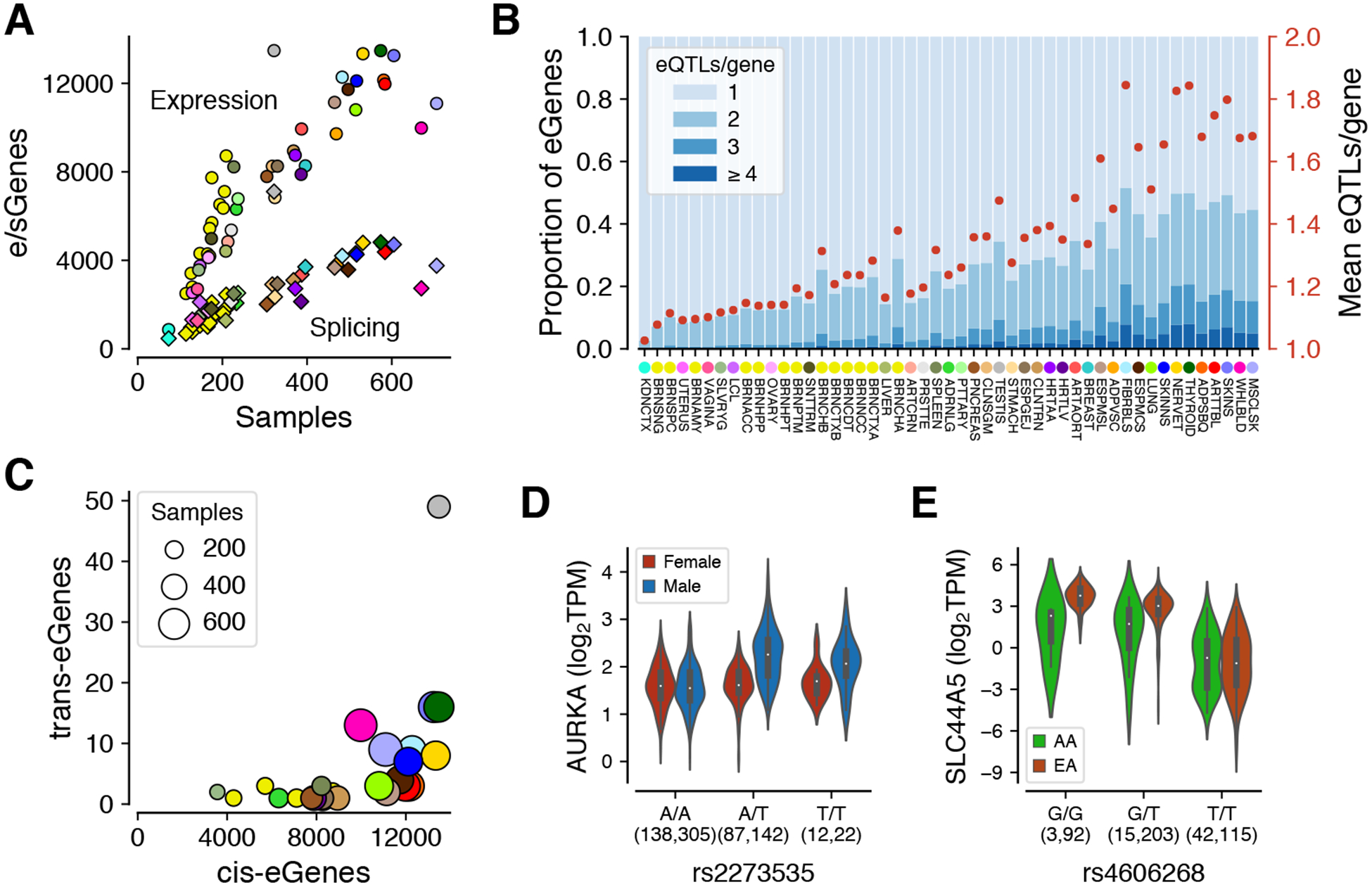
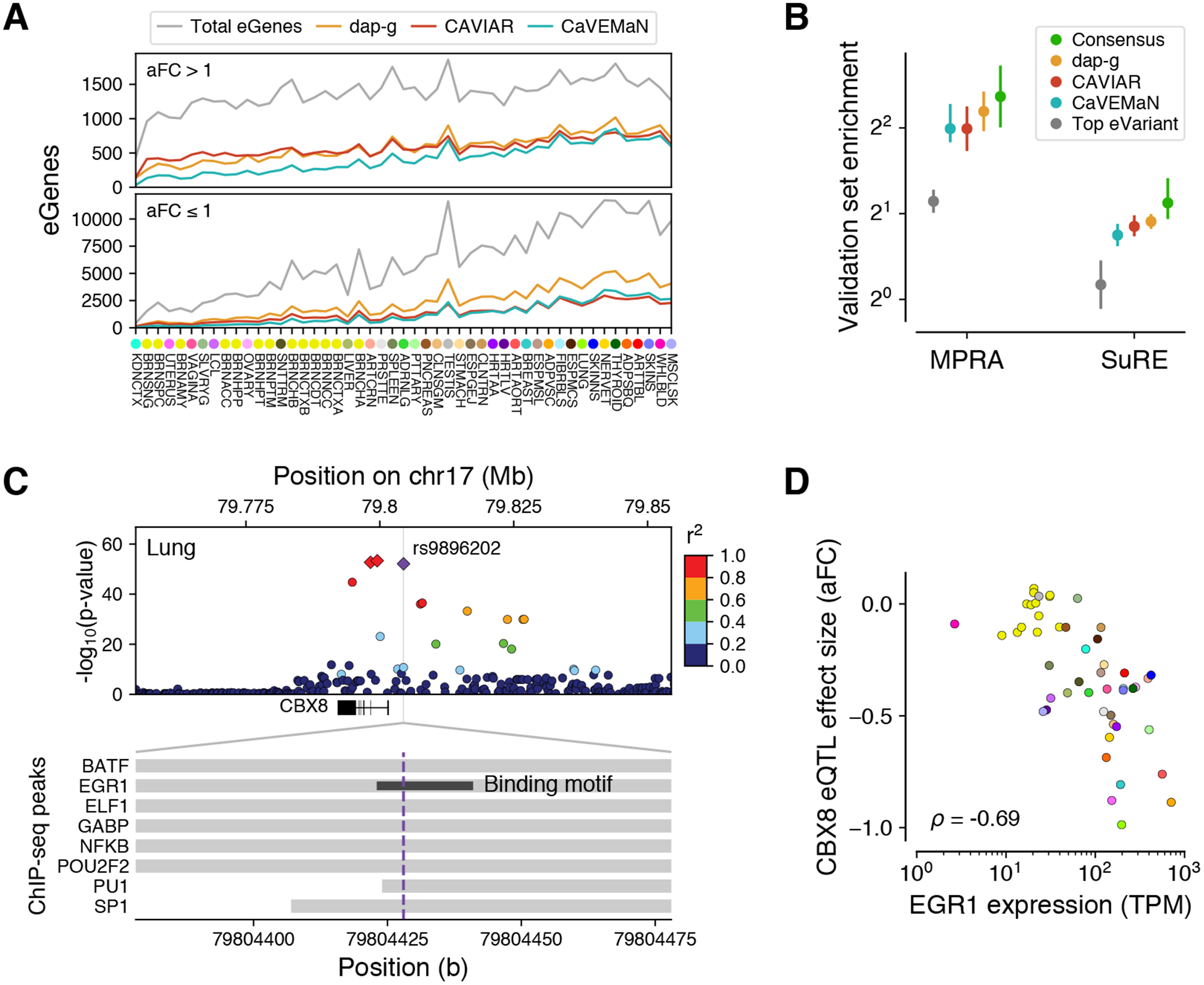
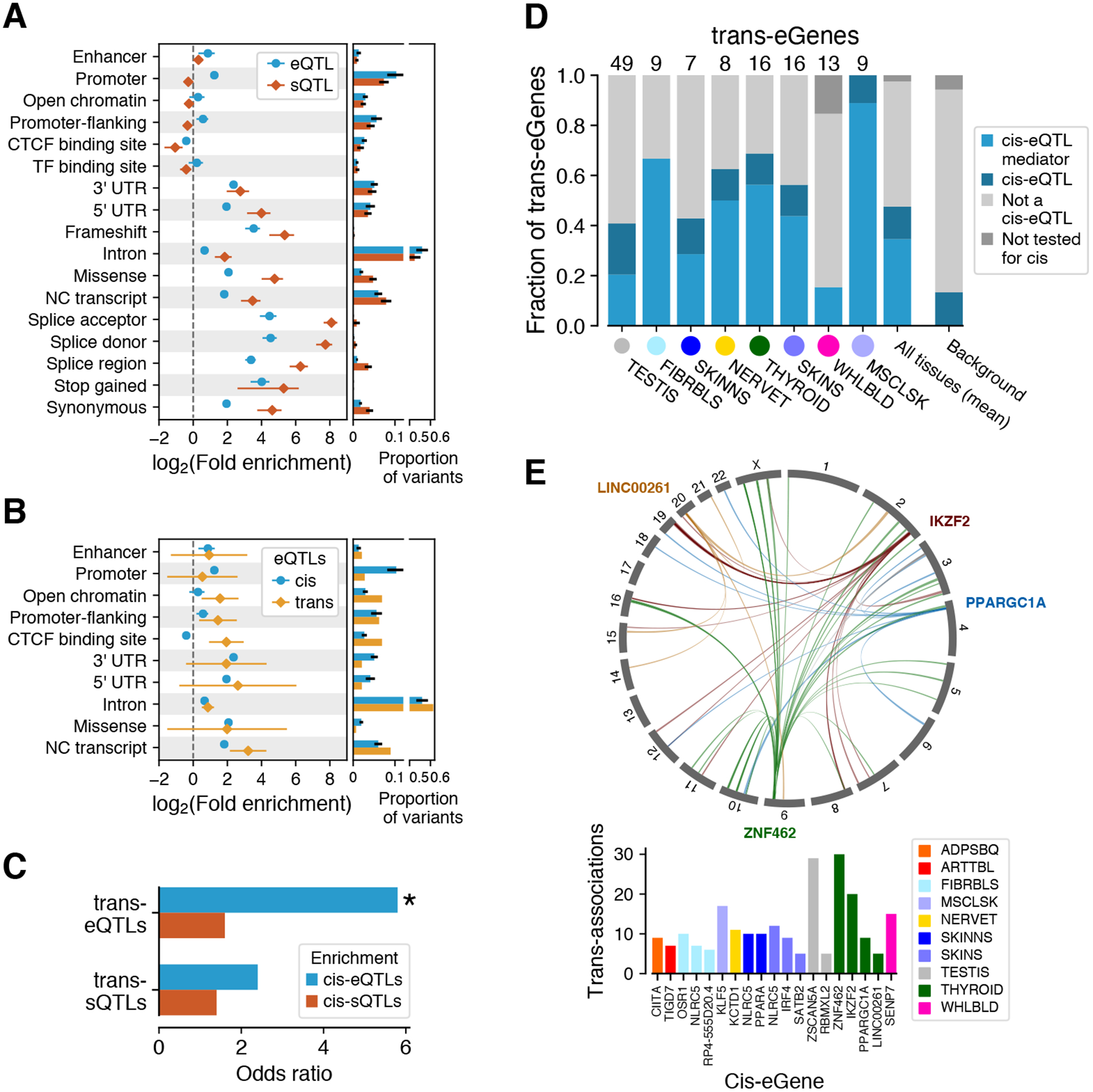
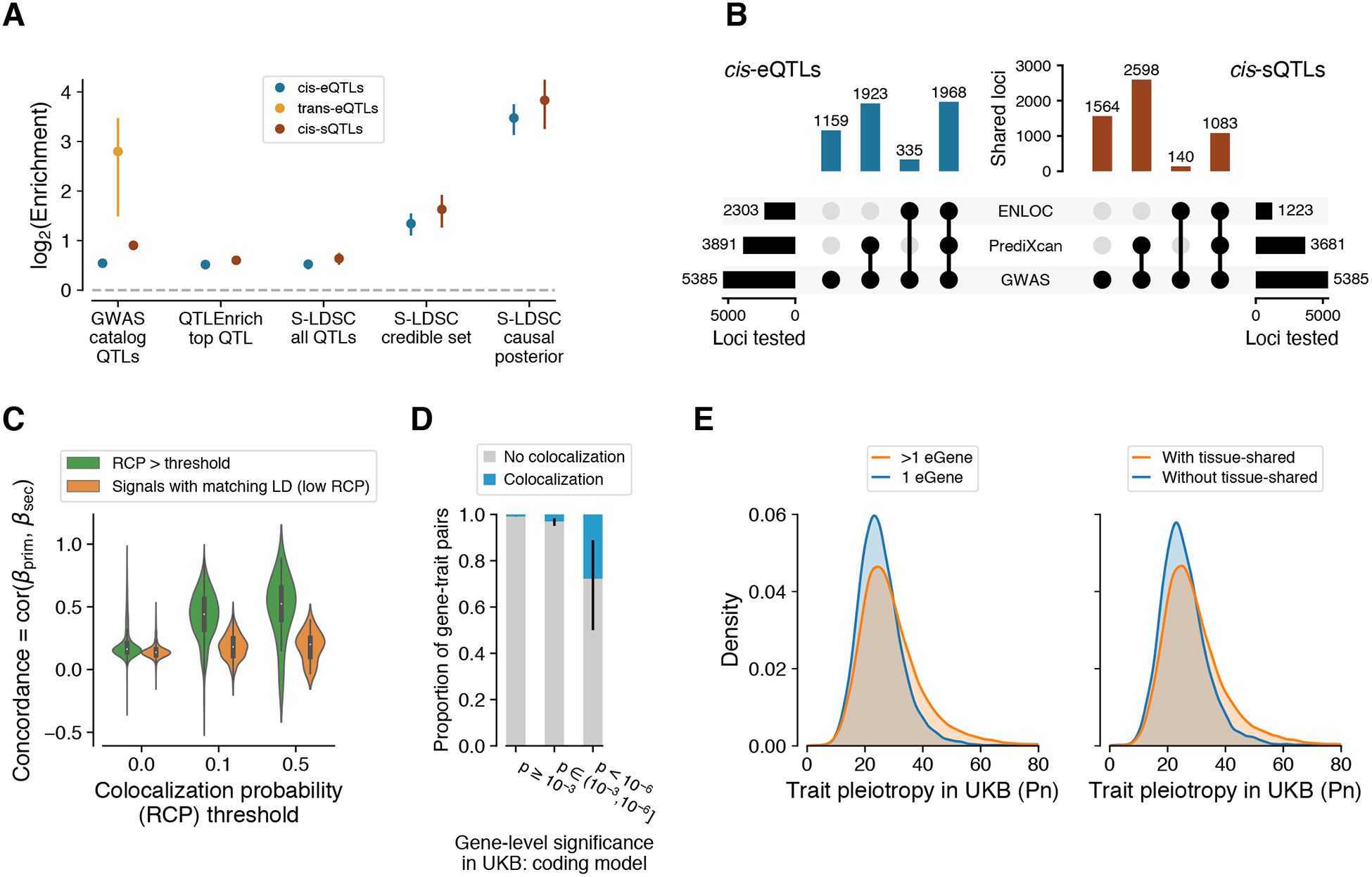
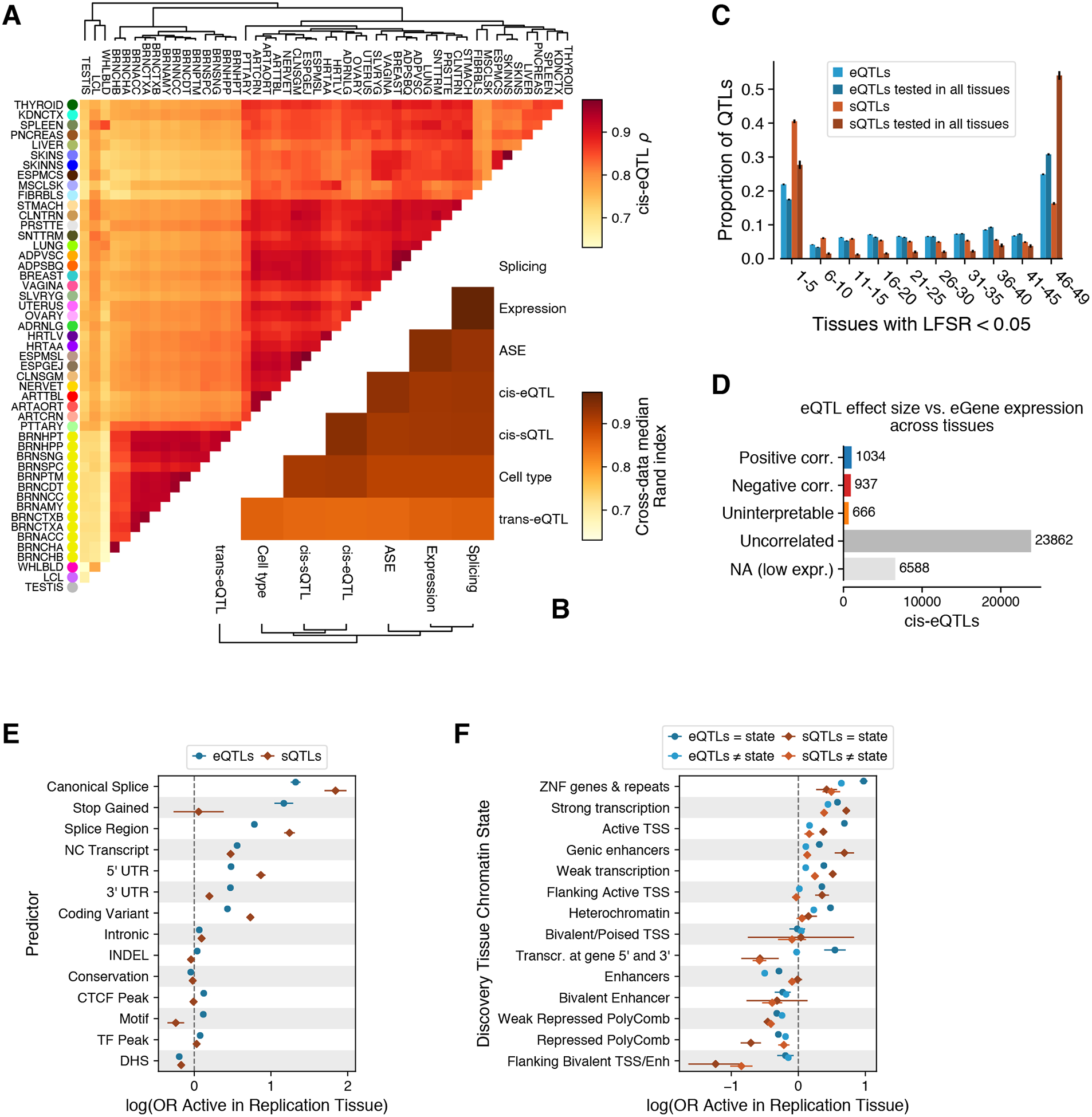
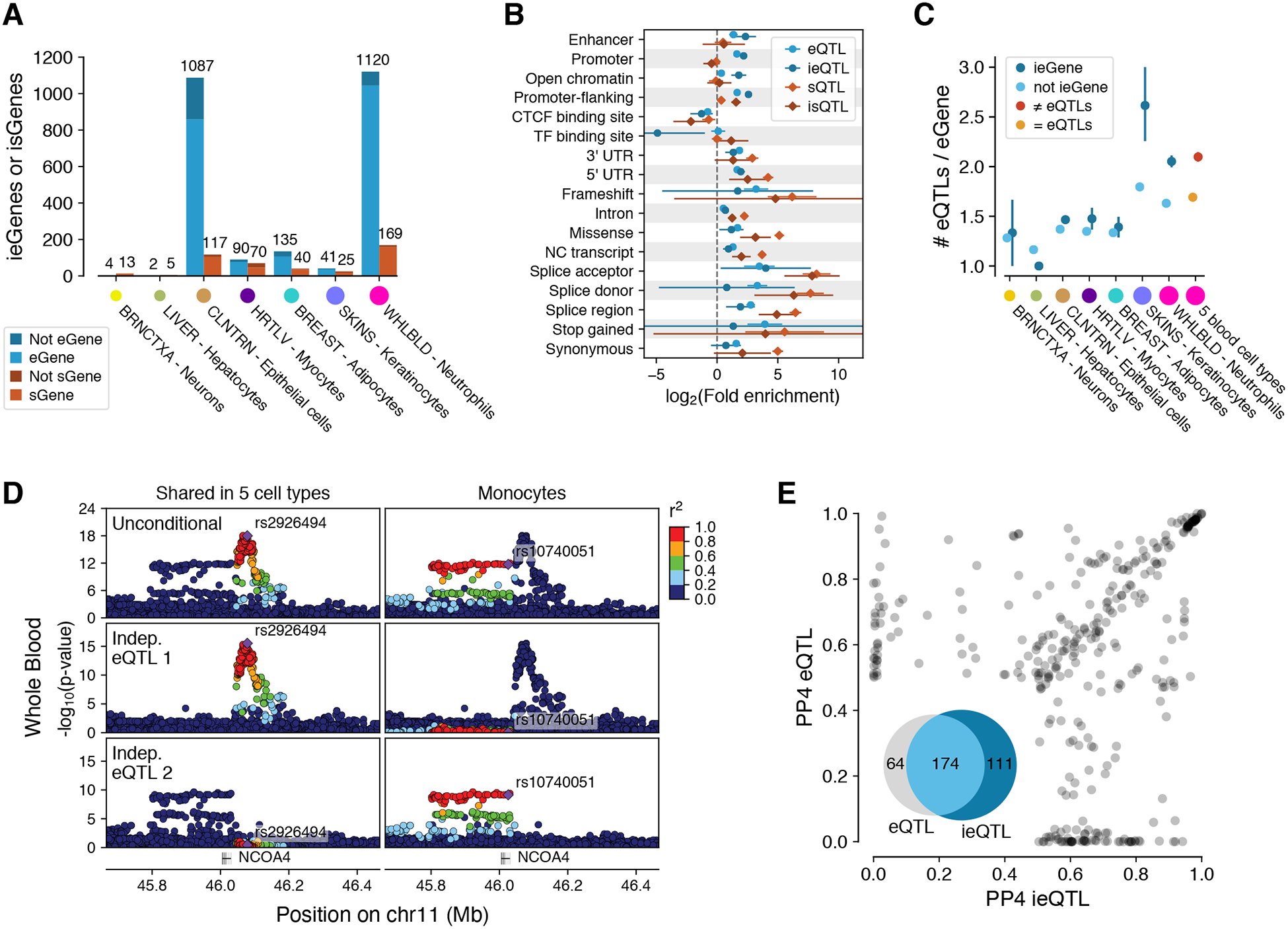
The Genotype-Tissue Expression (GTEx) project was established to characterize genetic effects on the transcriptome across human tissues and to link these regulatory mechanisms to trait and disease associations. Here, we present analyses of the version 8 data, examining 15,201 RNA-sequencing samples from 49 tissues of 838 postmortem donors. We comprehensively characterize genetic associations for gene expression and splicing in cis and trans, showing that regulatory associations are found for almost all genes, and describe the underlying molecular mechanisms and their contribution to allelic heterogeneity and pleiotropy of complex traits. Leveraging the large diversity of tissues, we provide insights into the tissue specificity of genetic effects and show that cell type composition is a key factor in understanding gene regulatory mechanisms in human tissues.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Competing interests
F.A. is an inventor on a patent application related to TensorQTL; S.E.C. is a co-founder, chief technology officer and stock owner at Variant Bio; E.R.G. is on the Editorial Board of Circulation Research, and does consulting for the City of Hope / Beckman Research Institut; E.T.D. is chairman and member of the board of Hybridstat LTD.; B.E.E. is on the scientific advisory boards of Celsius Therapeutics and Freenome; G.G. receives research funds from IBM and Pharmacyclics, and is an inventor on patent applications related to MuTect, ABSOLUTE, MutSig, MSMuTect, MSMutSig, POLYSOLVER and TensorQTL. G.G. is a founder, consultant and holds privately held equity in Scorpion Therapeutics; S.B.M. is on the scientific advisory board of MyOme; D.G.M. is a co-founder with equity in Goldfinch Bio, and has received research support from AbbVie, Astellas, Biogen, BioMarin, Eisai, Merck, Pfizer, and Sanofi-Genzyme; H.K.I. has received speaker honoraria from GSK and AbbVie.; T.L. is a scientific advisory board member of Variant Bio with equity and Goldfinch Bio. P.F. is member of the scientific advisory boards of Fabric Genomics, Inc., and Eagle Genomes, Ltd. P.G.F. is a partner of Bioinf2Bio.
Figures
Comment in
-
Reaching completion for GTEx.Nat Rev Genet. 2020 Dec;21(12):717. doi: 10.1038/s41576-020-00296-7. Nat Rev Genet. 2020. PMID: 33060849 No abstract available.
References
-
- Bonder MJ, et al. , Disease variants alter transcription factor levels and methylation of their binding sites. Nature Genetics 49, 131–138 (2017). - PubMed
Publication types
MeSH terms
Grants and funding
- R01 CA229618/CA/NCI NIH HHS/United States
- R01 MH107666/MH/NIMH NIH HHS/United States
- R01 MH106842/MH/NIMH NIH HHS/United States
- K99 HG009916/HG/NHGRI NIH HHS/United States
- R01 HL142028/HL/NHLBI NIH HHS/United States
- R01 HG006855/HG/NHGRI NIH HHS/United States
- R01 MH090951/MH/NIMH NIH HHS/United States
- UL1 TR002550/TR/NCATS NIH HHS/United States
- R01 HG011138/HG/NHGRI NIH HHS/United States
- U01 MH104393/MH/NIMH NIH HHS/United States
- R01 HG010480/HG/NHGRI NIH HHS/United States
- R01 DA006227/DA/NIDA NIH HHS/United States
- U41 HG002371/HG/NHGRI NIH HHS/United States
- R01 HG008150/HG/NHGRI NIH HHS/United States
- R01 MH109905/MH/NIMH NIH HHS/United States
- F31 HG010580/HG/NHGRI NIH HHS/United States
- UM1 HG008901/HG/NHGRI NIH HHS/United States
- U01 HG007593/HG/NHGRI NIH HHS/United States
- R01 GM124486/GM/NIGMS NIH HHS/United States
- R01 MH090936/MH/NIMH NIH HHS/United States
- U01 HG007598/HG/NHGRI NIH HHS/United States
- R01 AG057422/AG/NIA NIH HHS/United States
- T32 DK110919/DK/NIDDK NIH HHS/United States
- R01 HG002585/HG/NHGRI NIH HHS/United States
- T32 HG000044/HG/NHGRI NIH HHS/United States
- R01 HG010731/HG/NHGRI NIH HHS/United States
- R01 HL133218/HL/NHLBI NIH HHS/United States
- F32 HG009987/HG/NHGRI NIH HHS/United States
- R01 HG010067/HG/NHGRI NIH HHS/United States
- R01 MH090941/MH/NIMH NIH HHS/United States
- R01 GM122924/GM/NIGMS NIH HHS/United States
- U41 HG009494/HG/NHGRI NIH HHS/United States
- R01 MH101822/MH/NIMH NIH HHS/United States
- HHSN261200800001C/RC/CCR NIH HHS/United States
- R01 MH090937/MH/NIMH NIH HHS/United States
- UL1 TR001873/TR/NCATS NIH HHS/United States
- HHSN268201000029C/HL/NHLBI NIH HHS/United States
- HHSN261200800001E/CA/NCI NIH HHS/United States
- R01 MH101814/MH/NIMH NIH HHS/United States
- R35 HG010718/HG/NHGRI NIH HHS/United States
- P30 DK020595/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
