

CDK13 cooperates with CDK12 to control global RNA polymerase II processivity
- PMID: 32917631
- PMCID: PMC7190357
- DOI: 10.1126/sciadv.aaz5041
CDK13 cooperates with CDK12 to control global RNA polymerase II processivity
Abstract
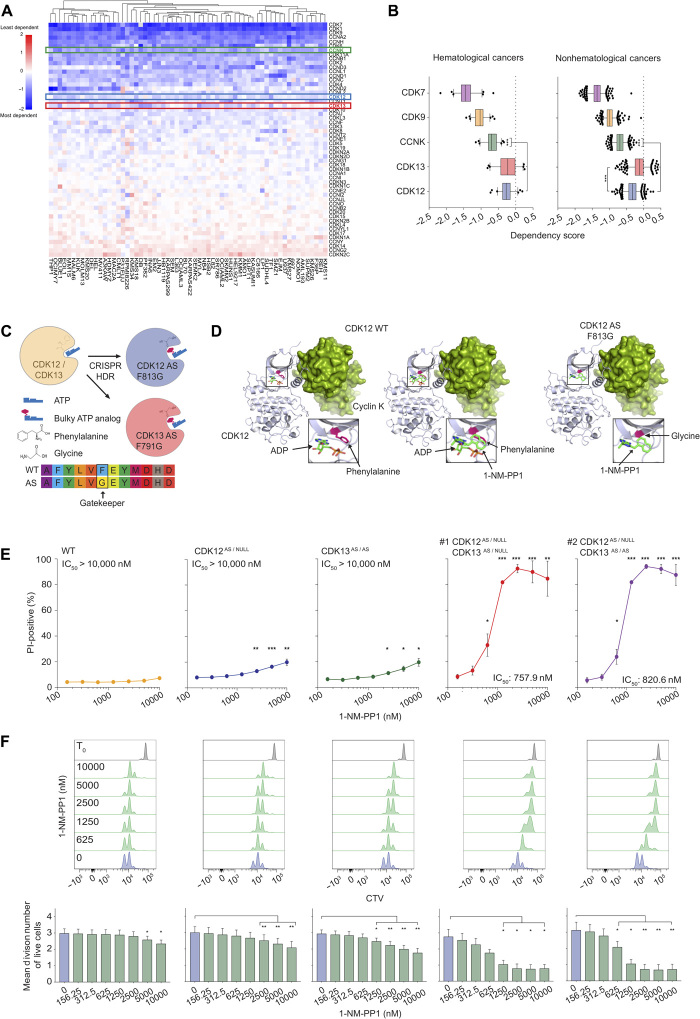
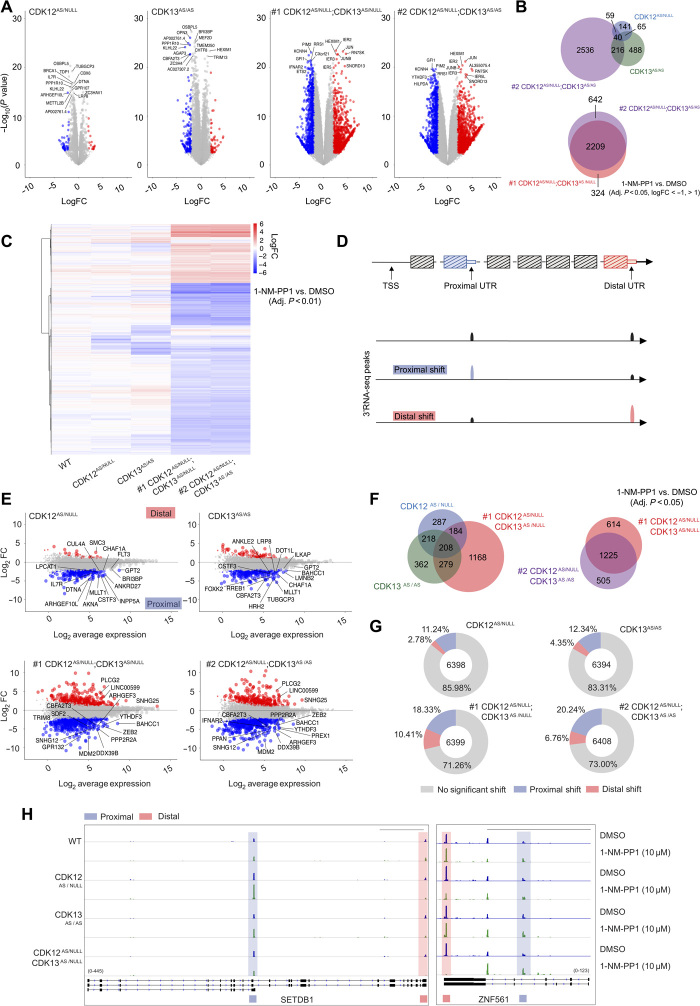
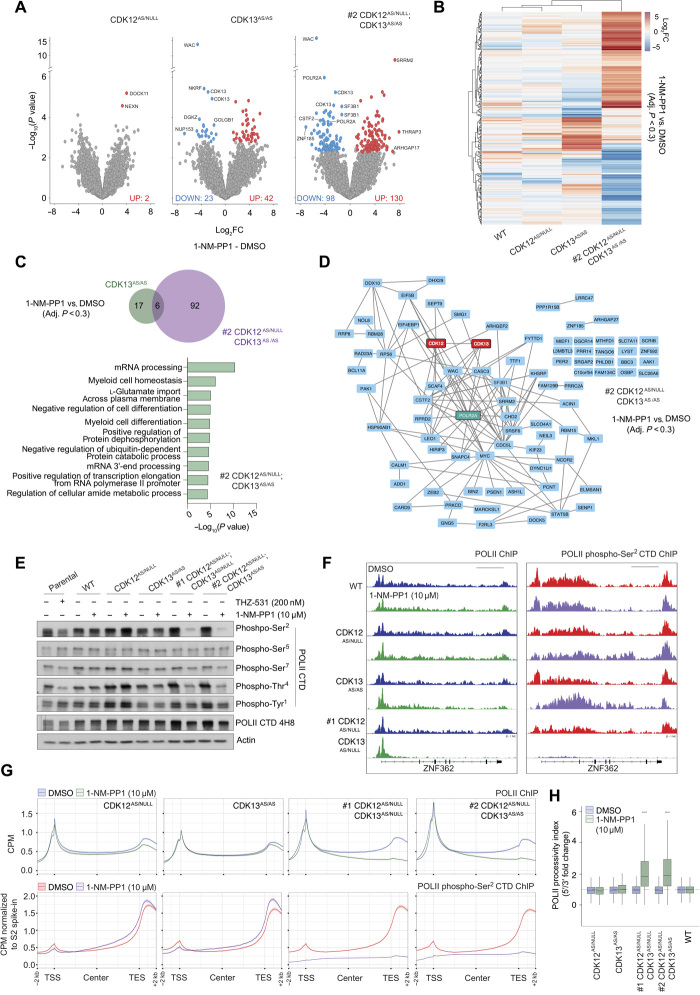
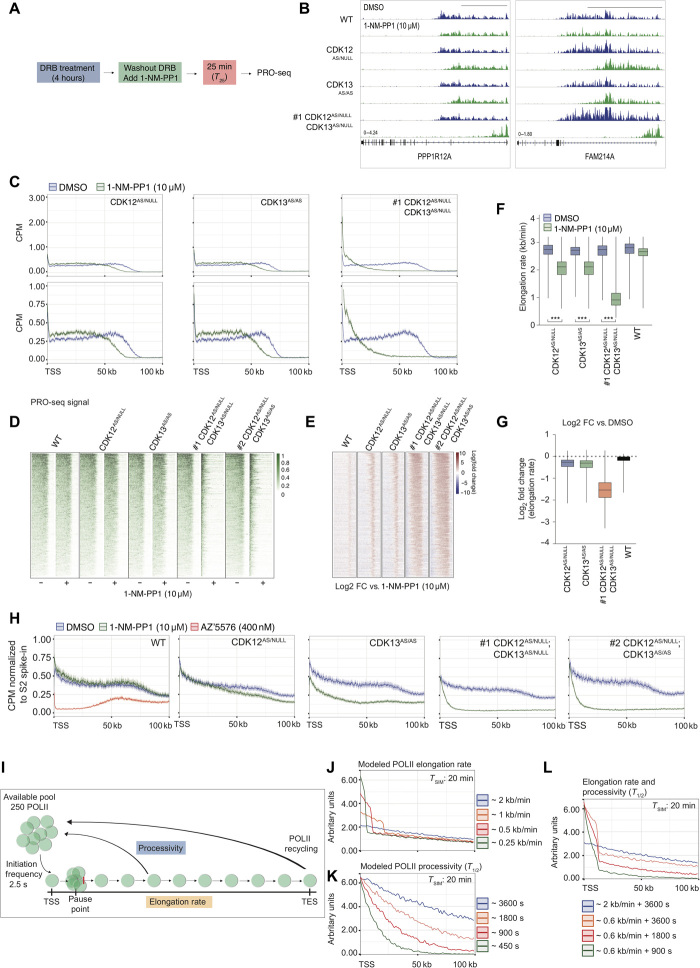
The RNA polymerase II (POLII)-driven transcription cycle is tightly regulated at distinct checkpoints by cyclin-dependent kinases (CDKs) and their cognate cyclins. The molecular events underpinning transcriptional elongation, processivity, and the CDK-cyclin pair(s) involved remain poorly understood. Using CRISPR-Cas9 homology-directed repair, we generated analog-sensitive kinase variants of CDK12 and CDK13 to probe their individual and shared biological and molecular roles. Single inhibition of CDK12 or CDK13 induced transcriptional responses associated with cellular growth signaling pathways and/or DNA damage, with minimal effects on cell viability. In contrast, dual kinase inhibition potently induced cell death, which was associated with extensive genome-wide transcriptional changes including widespread use of alternative 3' polyadenylation sites. At the molecular level, dual kinase inhibition resulted in the loss of POLII CTD phosphorylation and greatly reduced POLII elongation rates and processivity. These data define substantial redundancy between CDK12 and CDK13 and identify both as fundamental regulators of global POLII processivity and transcription elongation.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
References
-
- Chen F. X., Smith E. R., Shilatifard A., Born to run: Control of transcription elongation by RNA polymerase II. Nat. Rev. Mol. Cell Biol. 19, 464–478 (2018). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
