

Extracellular matrix stiffness determines DNA repair efficiency and cellular sensitivity to genotoxic agents
- PMID: 32917705
- PMCID: PMC7486107
- DOI: 10.1126/sciadv.abb2630
Extracellular matrix stiffness determines DNA repair efficiency and cellular sensitivity to genotoxic agents
Abstract
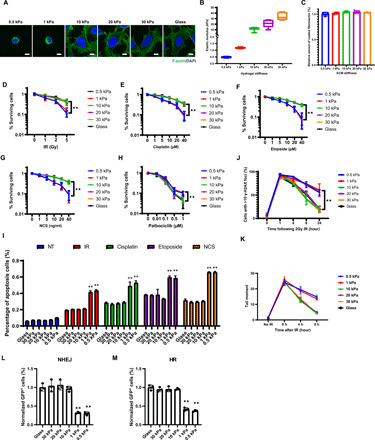
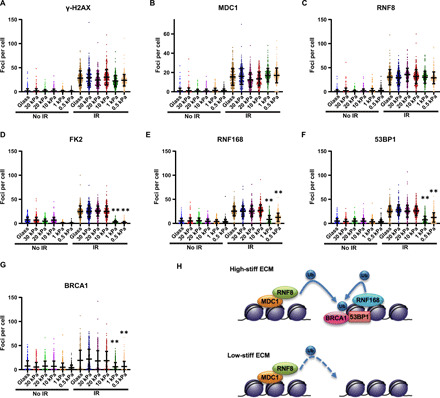
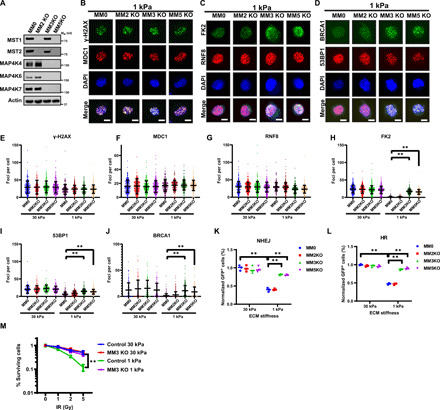
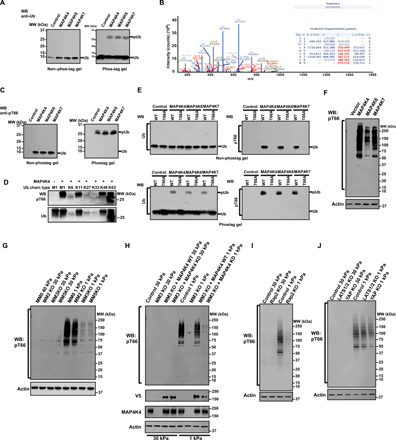
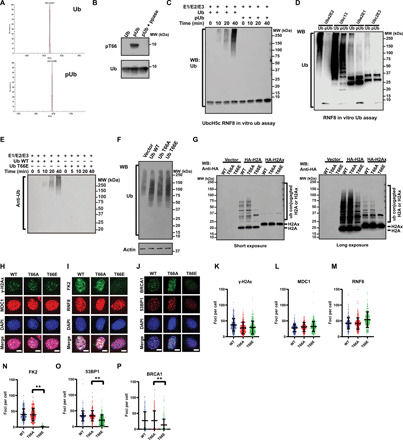
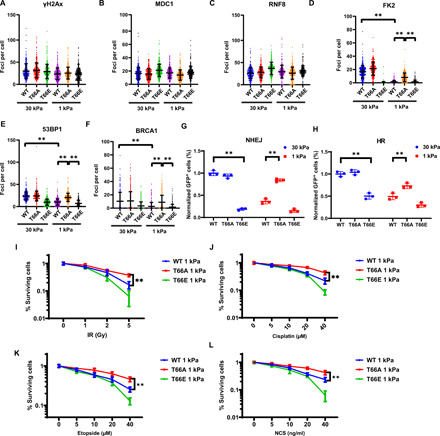
DNA double-strand breaks (DSBs) are highly toxic lesions that can drive genetic instability. These lesions also contribute to the efficacy of radiotherapy and many cancer chemotherapeutics. DNA repair efficiency is regulated by both intracellular and extracellular chemical signals. However, it is largely unknown whether this process is regulated by physical stimuli such as extracellular mechanical signals. Here, we report that DSB repair is regulated by extracellular mechanical signals. Low extracellular matrix (ECM) stiffness impairs DSB repair and renders cells sensitive to genotoxic agents. Mechanistically, we found that the MAP4K4/6/7 kinases are activated and phosphorylate ubiquitin in cells at low stiffness. Phosphorylated ubiquitin impairs RNF8-mediated ubiquitin signaling at DSB sites, leading to DSB repair deficiency. Our results thus demonstrate that ECM stiffness regulates DSB repair efficiency and genotoxic sensitivity through MAP4K4/6/7 kinase-mediated ubiquitin phosphorylation, providing a previously unidentified regulation in DSB-induced ubiquitin signaling.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
References
-
- Reinhardt H. C., Yaffe M. B., Phospho-Ser/Thr-binding domains: Navigating the cell cycle and DNA damage response. Nat. Rev. Mol. Cell Biol. 14, 563–580 (2013). - PubMed
-
- Bassing C. H., Chua K. F., Sekiguchi J., Suh H., Whitlow S. R., Fleming J. C., Monroe B. C., Ciccone D. N., Yan C., Vlasakova K., Livingston D. M., Ferguson D. O., Scully R., Alt F. W., Increased ionizing radiation sensitivity and genomic instability in the absence of histone H2AX. Proc. Natl. Acad. Sci. U.S.A. 99, 8173–8178 (2002). - PMC - PubMed
-
- Celeste A., Petersen S., Romanienko P. J., Fernandez-Capetillo O., Chen H. T., Sedelnikova O. A., Reina-San-Martin B., Coppola V., Meffre E., Difilippantonio M. J., Redon C., Pilch D. R., Olaru A., Eckhaus M., Camerini-Otero R. D., Tessarollo L., Livak F., Manova K., Bonner W. M., Nussenzweig M. C., Nussenzweig A., Genomic instability in mice lacking histone H2AX. Science 296, 922–927 (2002). - PMC - PubMed
-
- Rogakou E. P., Pilch D. R., Orr A. H., Ivanova V. S., Bonner W. M., DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J. Biol. Chem. 273, 5858–5868 (1998). - PubMed
-
- Stewart G. S., Wang B., Bignell C. R., Taylor A. M. R., Elledge S. J., MDC1 is a mediator of the mammalian DNA damage checkpoint. Nature 421, 961–966 (2003). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
