

Wnt/β-catenin signaling regulates adipose tissue lipogenesis and adipocyte-specific loss is rigorously defended by neighboring stromal-vascular cells
- PMID: 32919095
- PMCID: PMC7554252
- DOI: 10.1016/j.molmet.2020.101078
Wnt/β-catenin signaling regulates adipose tissue lipogenesis and adipocyte-specific loss is rigorously defended by neighboring stromal-vascular cells
Abstract
Objective: Canonical Wnt/β-catenin signaling is a well-studied endogenous regulator of mesenchymal cell fate determination, promoting osteoblastogenesis and inhibiting adipogenesis. However, emerging genetic evidence in humans links a number of Wnt pathway members to body fat distribution, obesity, and metabolic dysfunction, suggesting that this pathway also functions in adipocytes. Recent studies in mice have uncovered compelling evidence that the Wnt signaling pathway plays important roles in adipocyte metabolism, particularly under obesogenic conditions. However, complexities in Wnt signaling and differences in experimental models and approaches have thus far limited our understanding of its specific roles in this context.
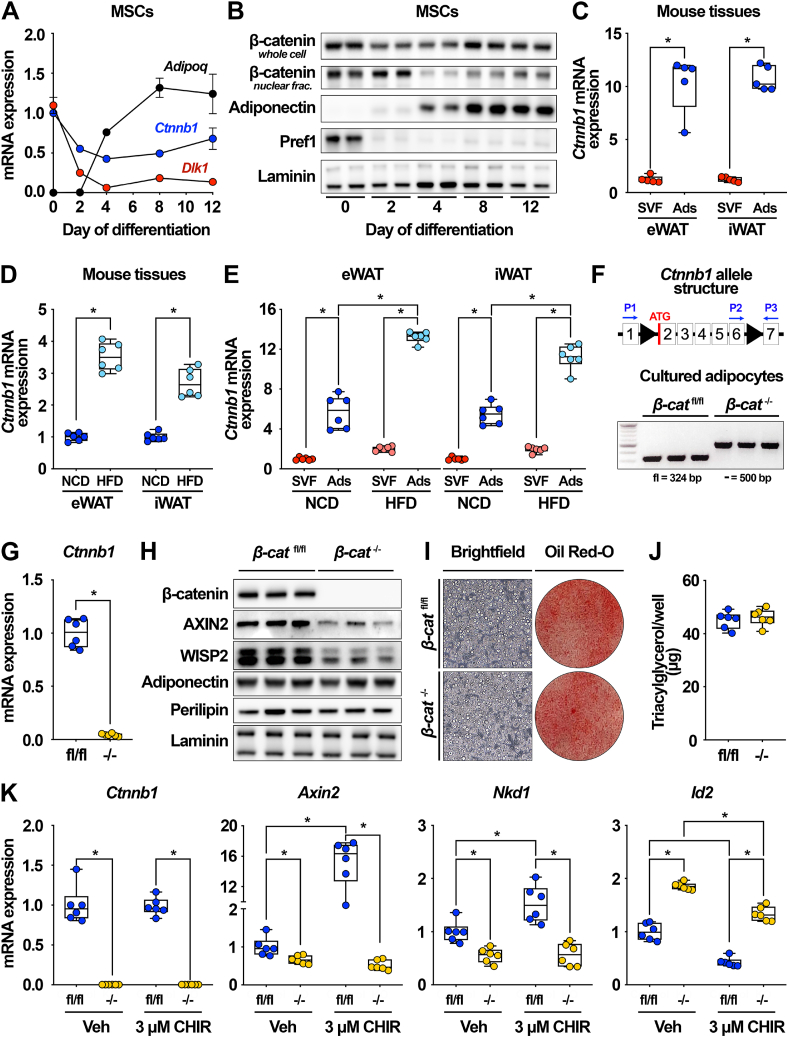
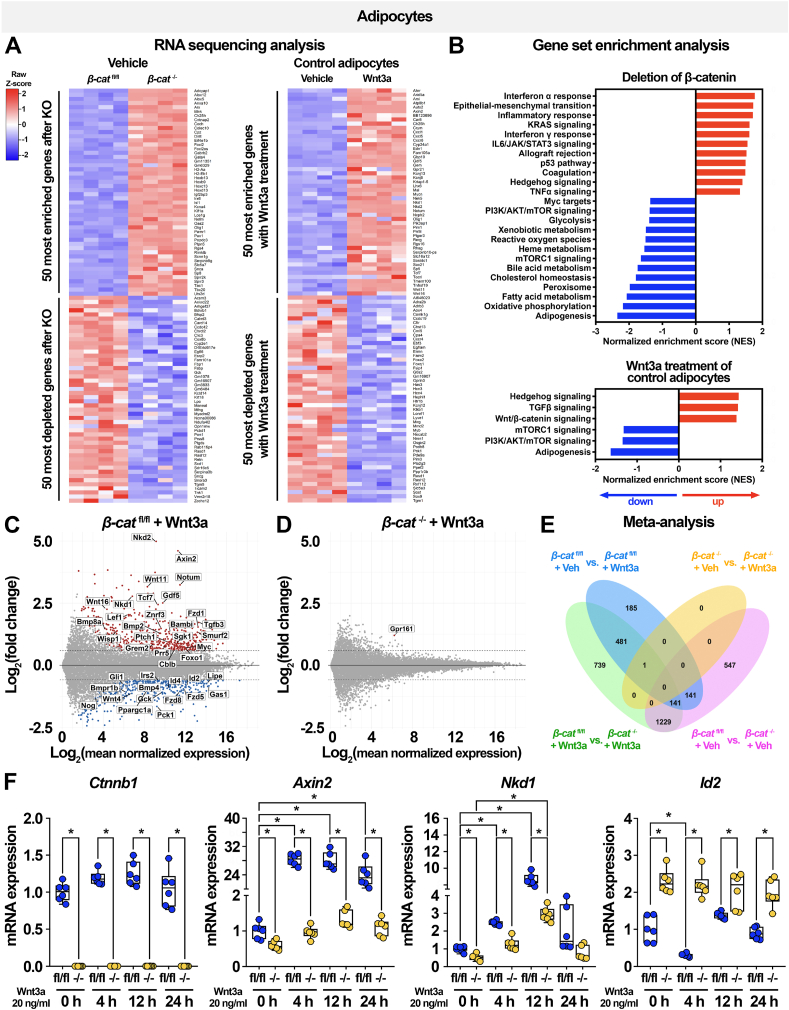
Methods: To investigate roles of the canonical Wnt pathway in the regulation of adipocyte metabolism, we generated adipocyte-specific β-catenin (β-cat) knockout mouse and cultured cell models. We used RNA sequencing, ChIP sequencing, and molecular approaches to assess expression of Wnt targets and lipogenic genes. We then used functional assays to evaluate effects of β-catenin deficiency on adipocyte metabolism, including lipid and carbohydrate handling. In mice maintained on normal chow and high-fat diets, we assessed the cellular and functional consequences of adipocyte-specific β-catenin deletion on adipose tissues and systemic metabolism.
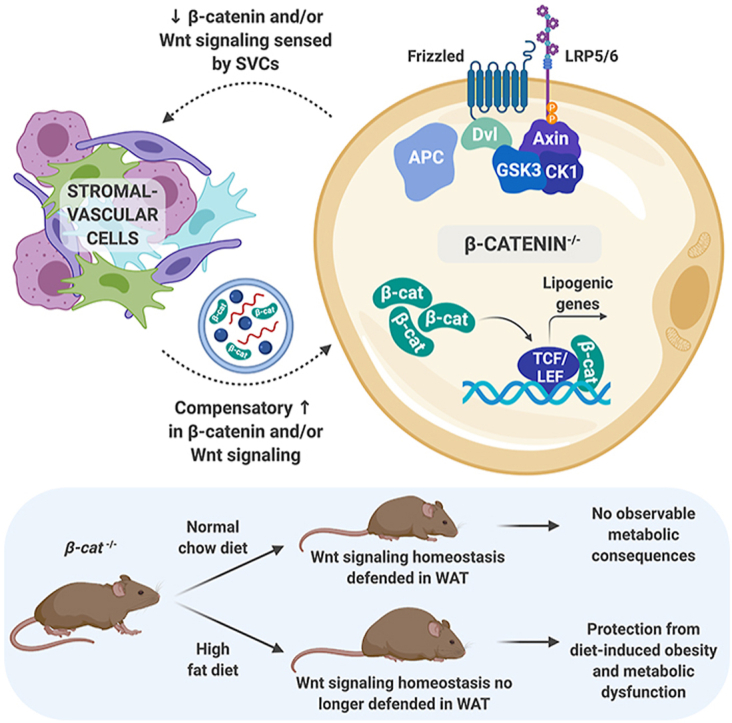
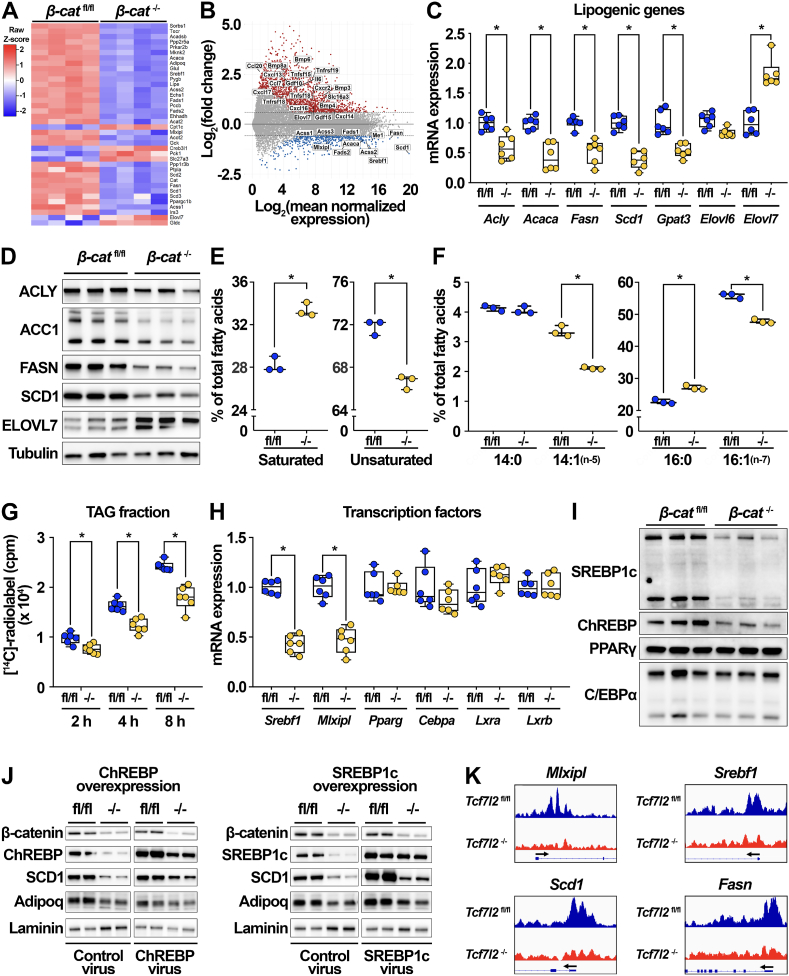
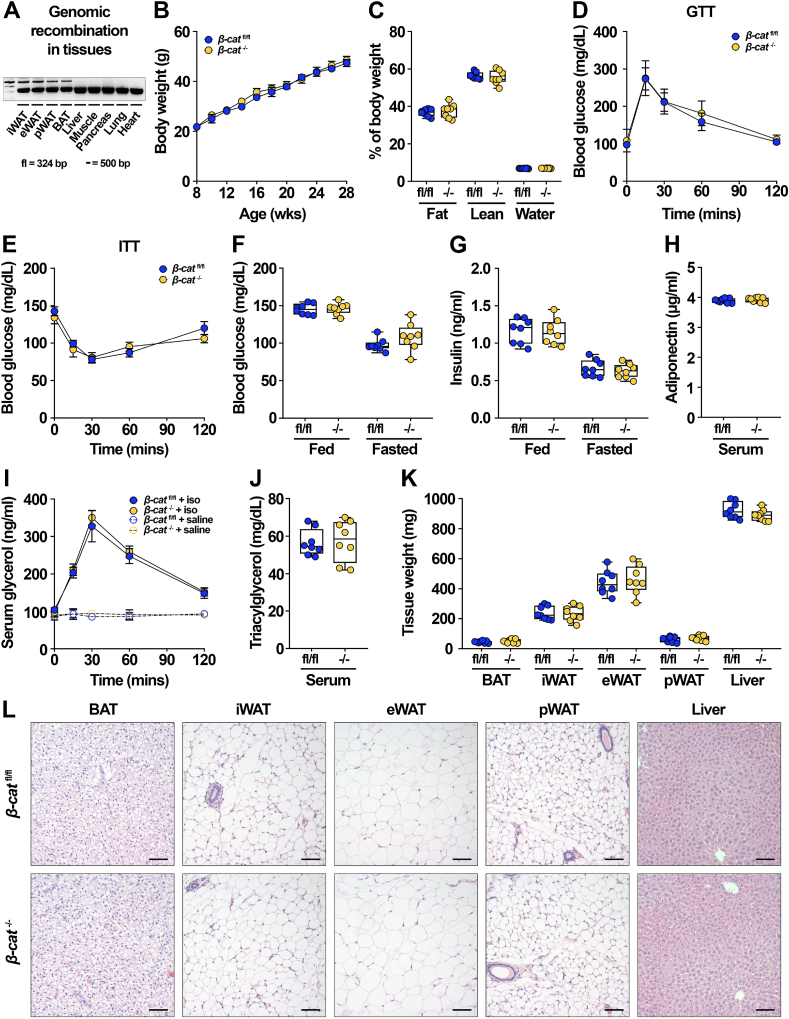
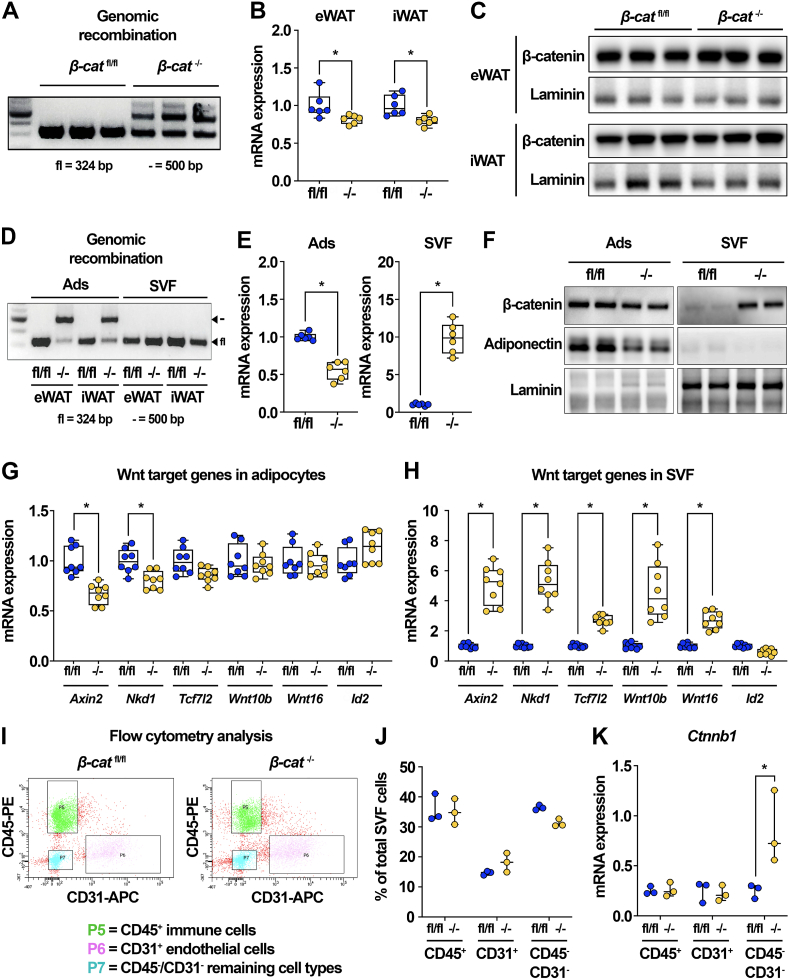
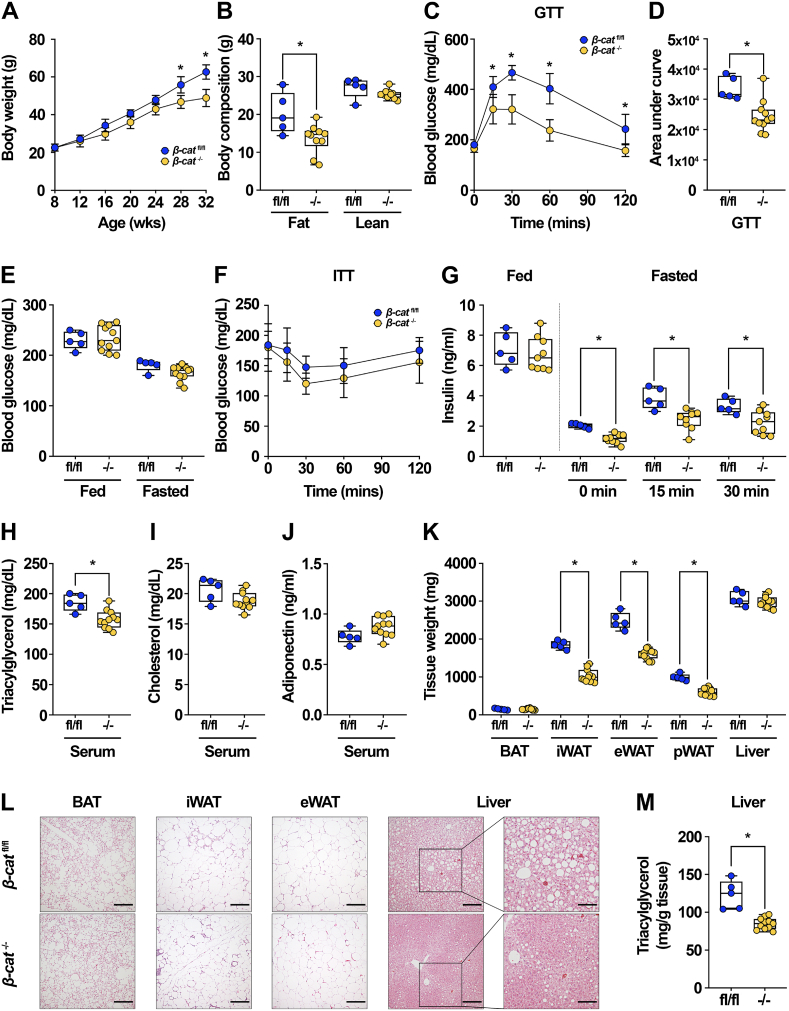
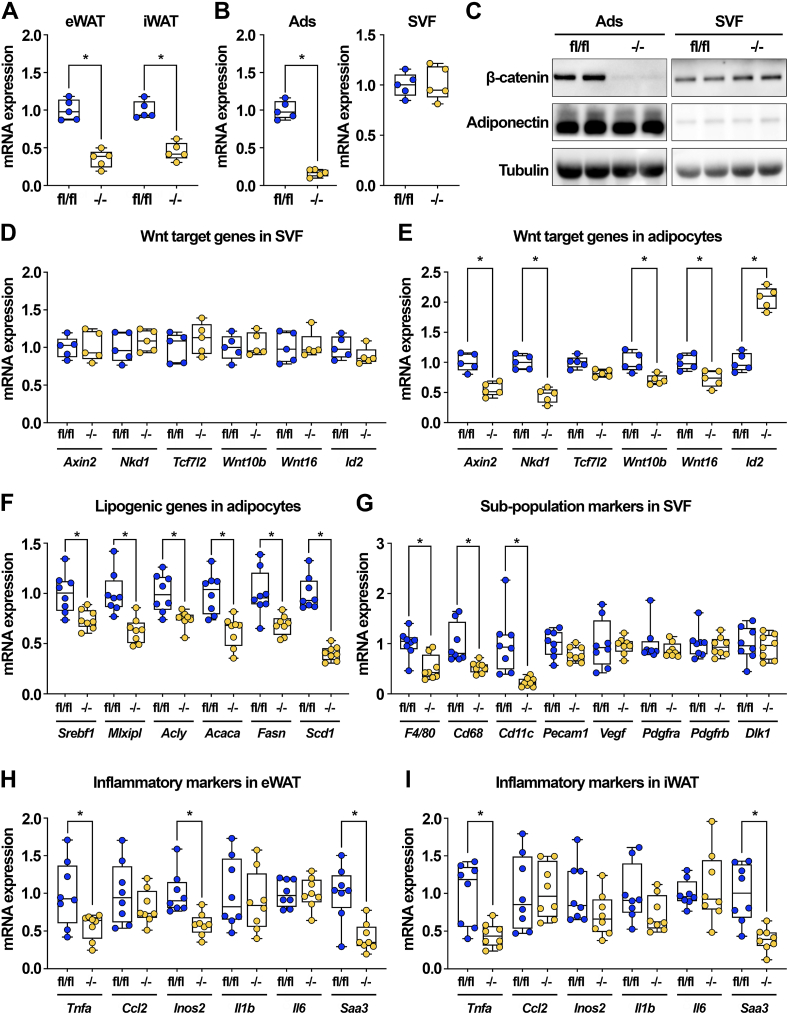
Results: We report that in adipocytes, the canonical Wnt/β-catenin pathway regulates de novo lipogenesis (DNL) and fatty acid monounsaturation. Further, β-catenin mediates effects of Wnt signaling on lipid metabolism in part by transcriptional regulation of Mlxipl and Srebf1. Intriguingly, adipocyte-specific loss of β-catenin is sensed and defended by CD45-/CD31- stromal cells to maintain tissue-wide Wnt signaling homeostasis in chow-fed mice. With long-term high-fat diet, this compensatory mechanism is overridden, revealing that β-catenin deletion promotes resistance to diet-induced obesity and adipocyte hypertrophy and subsequent protection from metabolic dysfunction.
Conclusions: Taken together, our studies demonstrate that Wnt signaling in adipocytes is required for lipogenic gene expression, de novo lipogenesis, and lipid desaturation. In addition, adipose tissues rigorously defend Wnt signaling homeostasis under standard nutritional conditions, such that stromal-vascular cells sense and compensate for adipocyte-specific loss. These findings underscore the critical importance of this pathway in adipocyte lipid metabolism and adipose tissue function.
Keywords: Adipocyte; Adipose tissue; Lipogenesis; Metabolism; Wnt signaling; β-catenin.
Copyright © 2020 The Author(s). Published by Elsevier GmbH.. All rights reserved.
Figures
References
-
- Cadigan K.M., Nusse R. Wnt signaling: a common theme in animal development. Genes & Development. 1997;11(24):3286–3305. - PubMed
-
- Clevers H. Wnt/beta-catenin signaling in development and disease. Cell. 2006;127(3):469–480. - PubMed
-
- Logan C.Y., Nusse R. The Wnt signaling pathway in development and disease. Annual Review of Cell and Developmental Biology. 2004;20:781–810. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R24 DK092759/DK/NIDDK NIH HHS/United States
- R01 DK125513/DK/NIDDK NIH HHS/United States
- R24 DK084970/DK/NIDDK NIH HHS/United States
- T32 DK101357/DK/NIDDK NIH HHS/United States
- T32 GM007863/GM/NIGMS NIH HHS/United States
- N02 DK062876/DK/NIDDK NIH HHS/United States
- R01 DK062876/DK/NIDDK NIH HHS/United States
- P30 DK020572/DK/NIDDK NIH HHS/United States
- U24 DK097153/DK/NIDDK NIH HHS/United States
- R01 DK121759/DK/NIDDK NIH HHS/United States
- U2C DK110768/DK/NIDDK NIH HHS/United States
- R56 DK062876/DK/NIDDK NIH HHS/United States
- P30 DK089503/DK/NIDDK NIH HHS/United States
- T32 HD007505/HD/NICHD NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
