

Vitamin E treatment in NAFLD patients demonstrates that oxidative stress drives steatosis through upregulation of de-novo lipogenesis
- PMID: 32920226
- PMCID: PMC7494510
- DOI: 10.1016/j.redox.2020.101710
Vitamin E treatment in NAFLD patients demonstrates that oxidative stress drives steatosis through upregulation of de-novo lipogenesis
Abstract
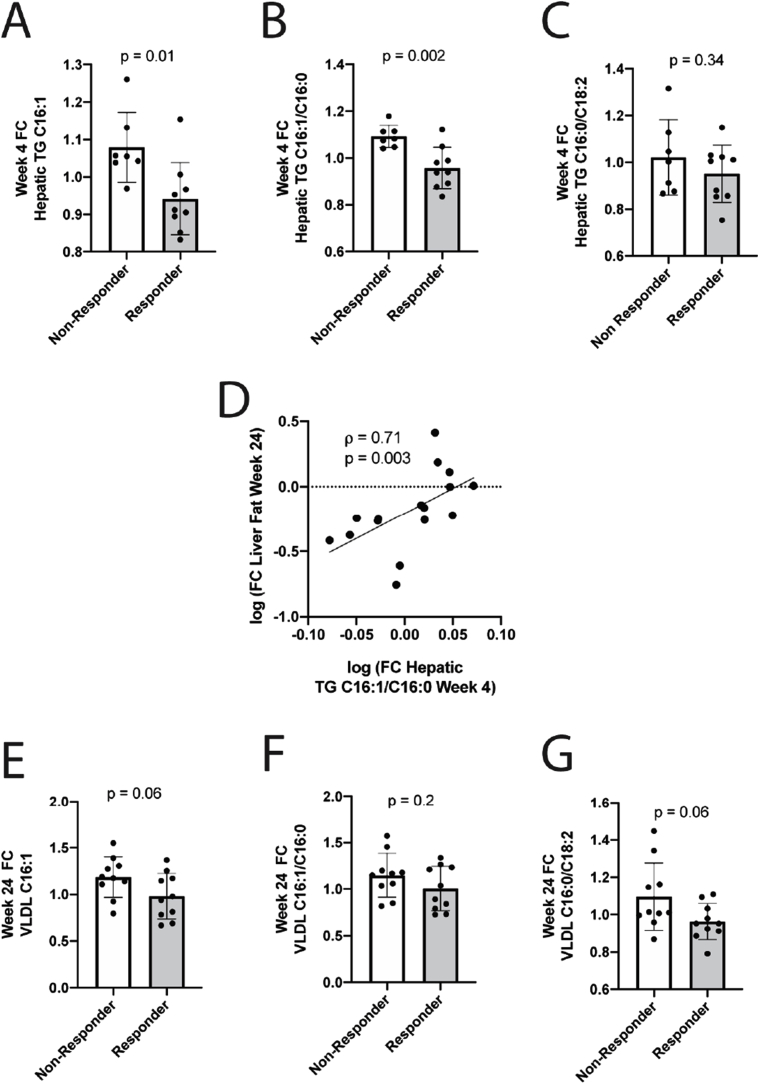
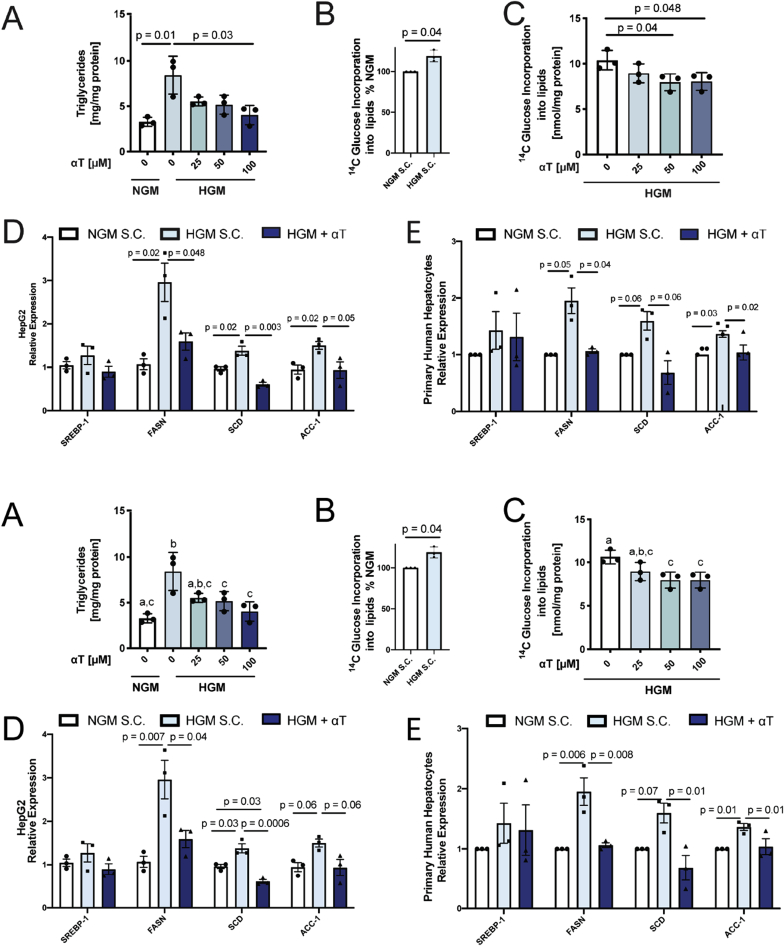
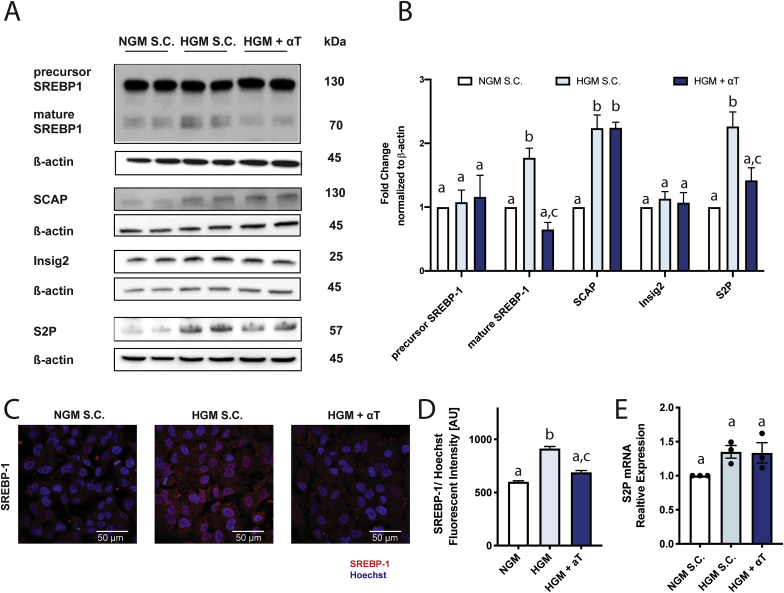
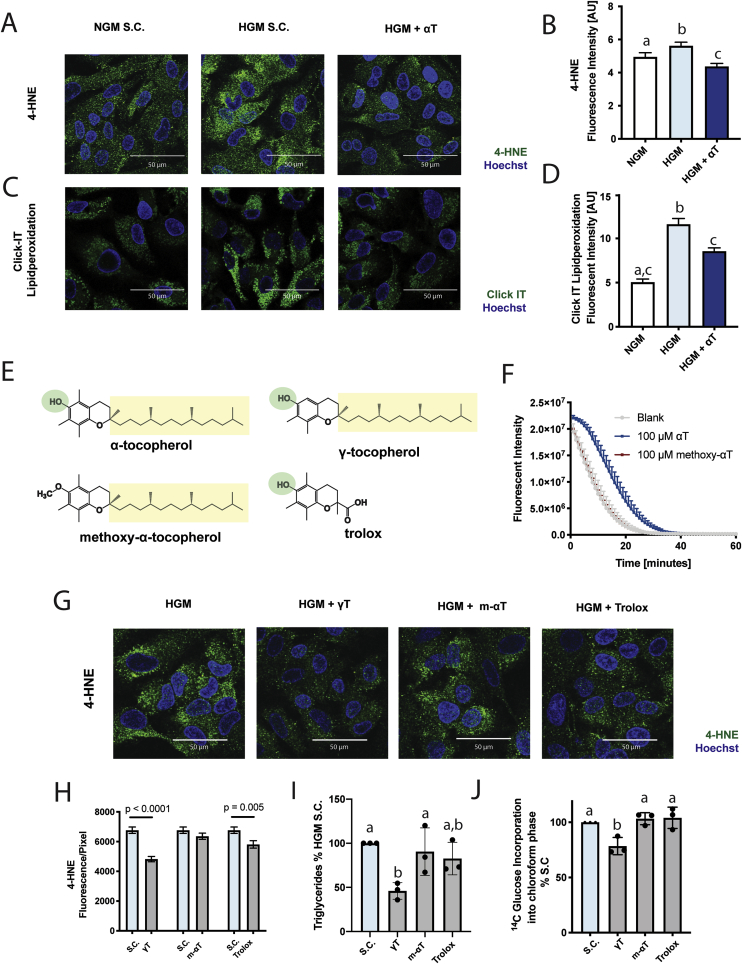
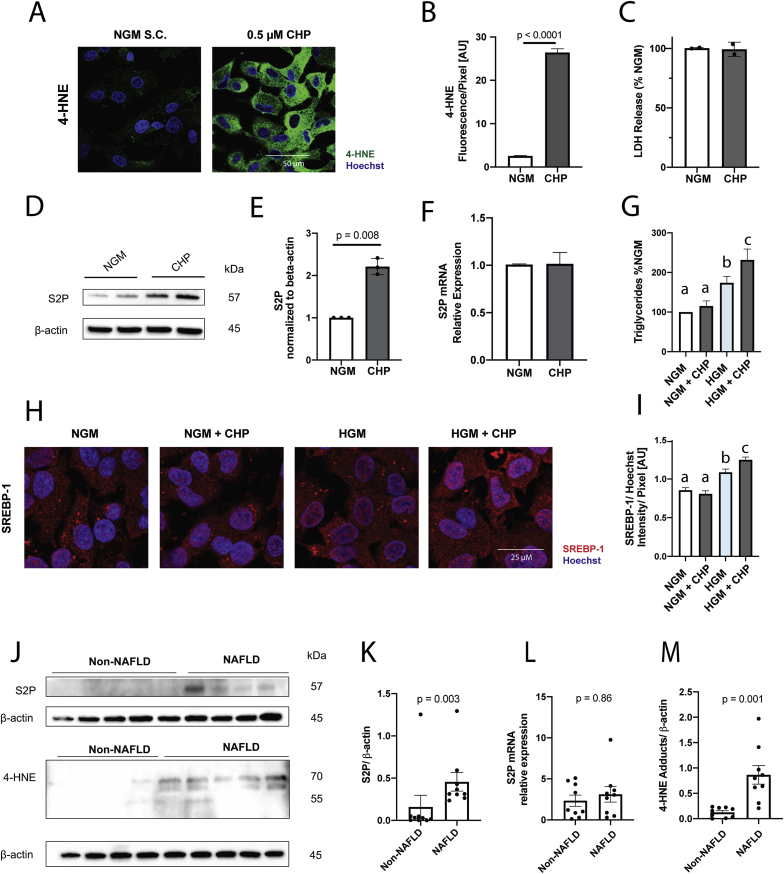
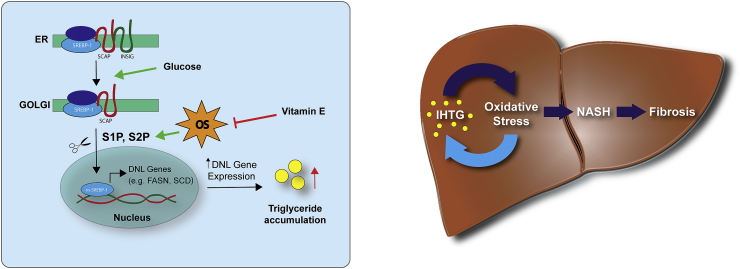
Oxidative stress (OS) in non-alcoholic fatty liver disease (NAFLD) promotes liver injury and inflammation. Treatment with vitamin E (α-tocopherol, αT), a lipid-soluble antioxidant, improves liver injury but also decreases steatosis, thought to be upstream of OS, through an unknown mechanism. To elucidate the mechanism, we combined a mechanistic human trial interrogating pathways of intrahepatic triglyceride (IHTG) accumulation and in vitro experiments. 50% of NAFLD patients (n = 20) treated with αT (200-800 IU/d) for 24 weeks had a ≥ 25% relative decrease in IHTG by magnetic resonance spectroscopy. Paired liver biopsies at baseline and week 4 of treatment revealed a decrease in markers of hepatic de novo lipogenesis (DNL) that strongly predicted week 24 response. In vitro, using HepG2 cells and primary human hepatocytes, αT inhibited glucose-induced DNL by decreasing SREBP-1 processing and lipogenic gene expression. This mechanism is dependent on the antioxidant capacity of αT, as redox-silenced methoxy-αT is unable to inhibit DNL in vitro. OS by itself was sufficient to increase S2P expression in vitro, and S2P is upregulated in NAFLD livers. In summary, we utilized αT to demonstrate a vicious cycle in which NAFLD generates OS, which feeds back to augment DNL and increases steatosis. Clinicaltrials.gov: NCT01792115.
Keywords: NAFLD; Non-alcoholic fatty liver disease; Oxidative stress; S1P; S2P; Vitamin E; de novo lipogenesis.
Published by Elsevier B.V.
Conflict of interest statement
The authors have declared no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
