

Novel blood coagulation molecules: Skeletal muscle myosin and cardiac myosin
- PMID: 32920971
- PMCID: PMC7819347
- DOI: 10.1111/jth.15097
Novel blood coagulation molecules: Skeletal muscle myosin and cardiac myosin
Abstract
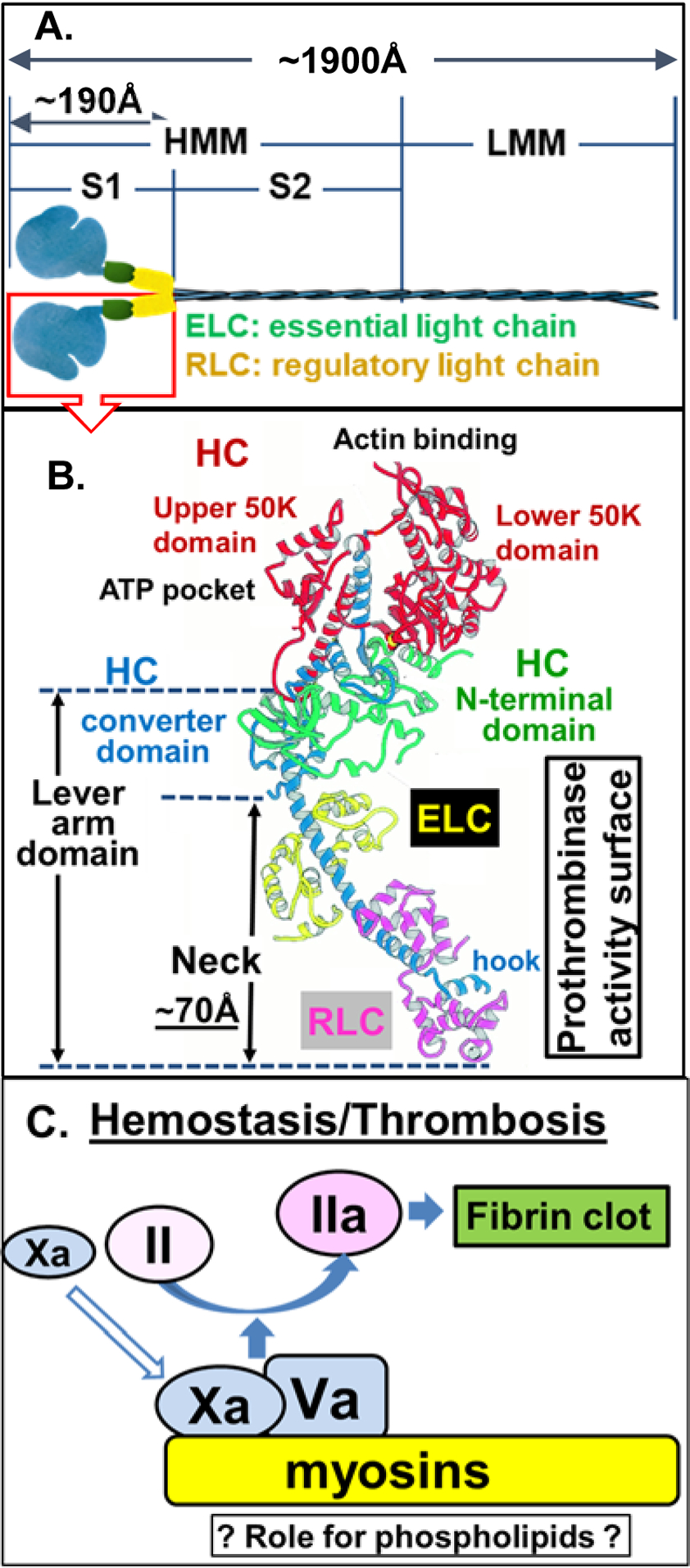
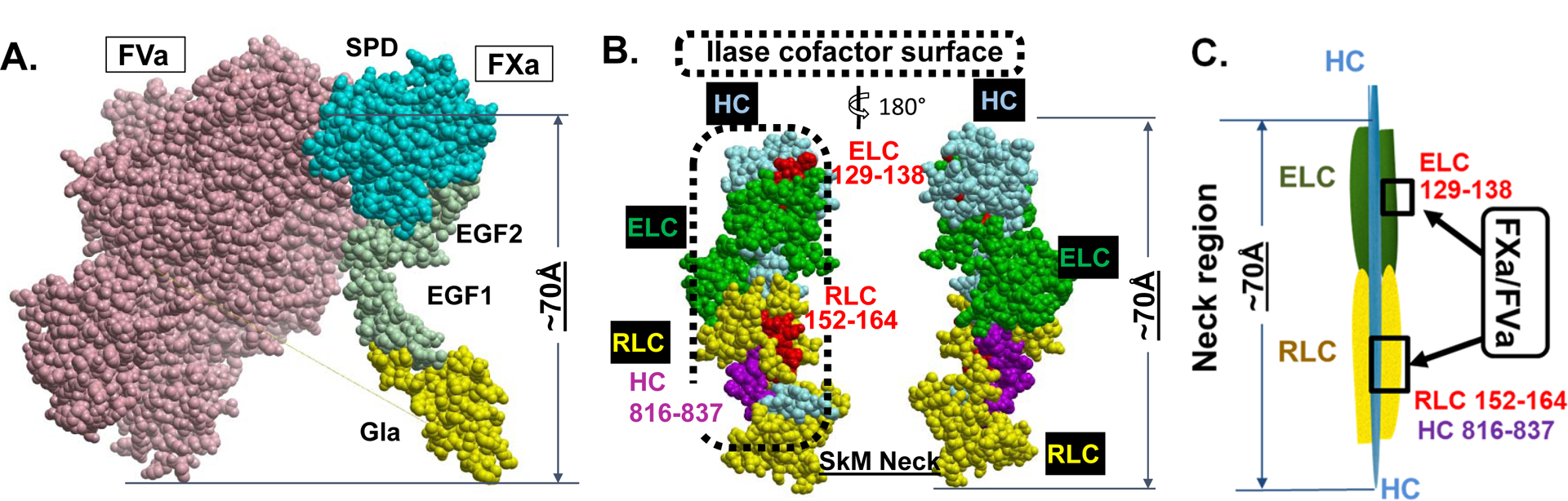
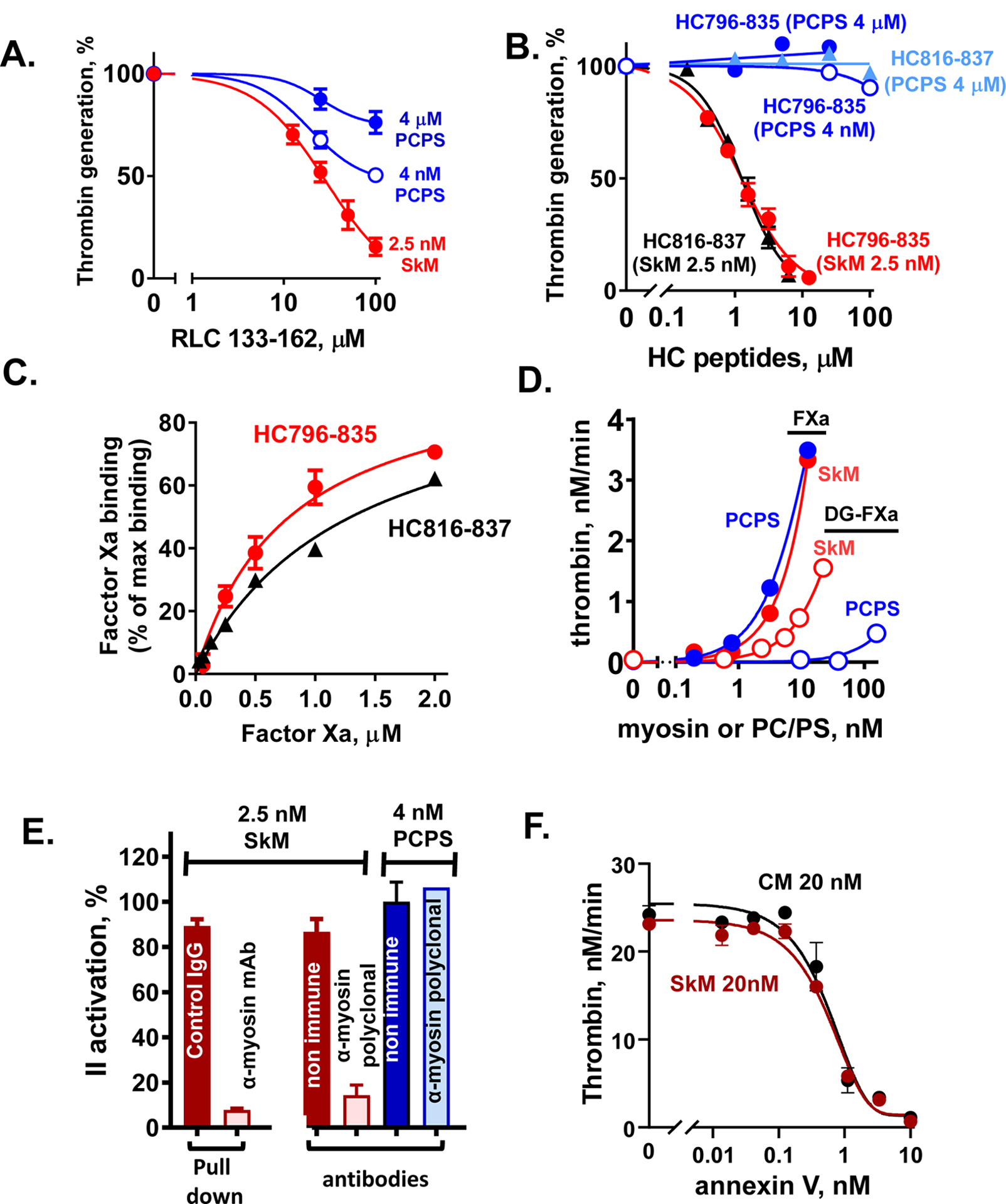
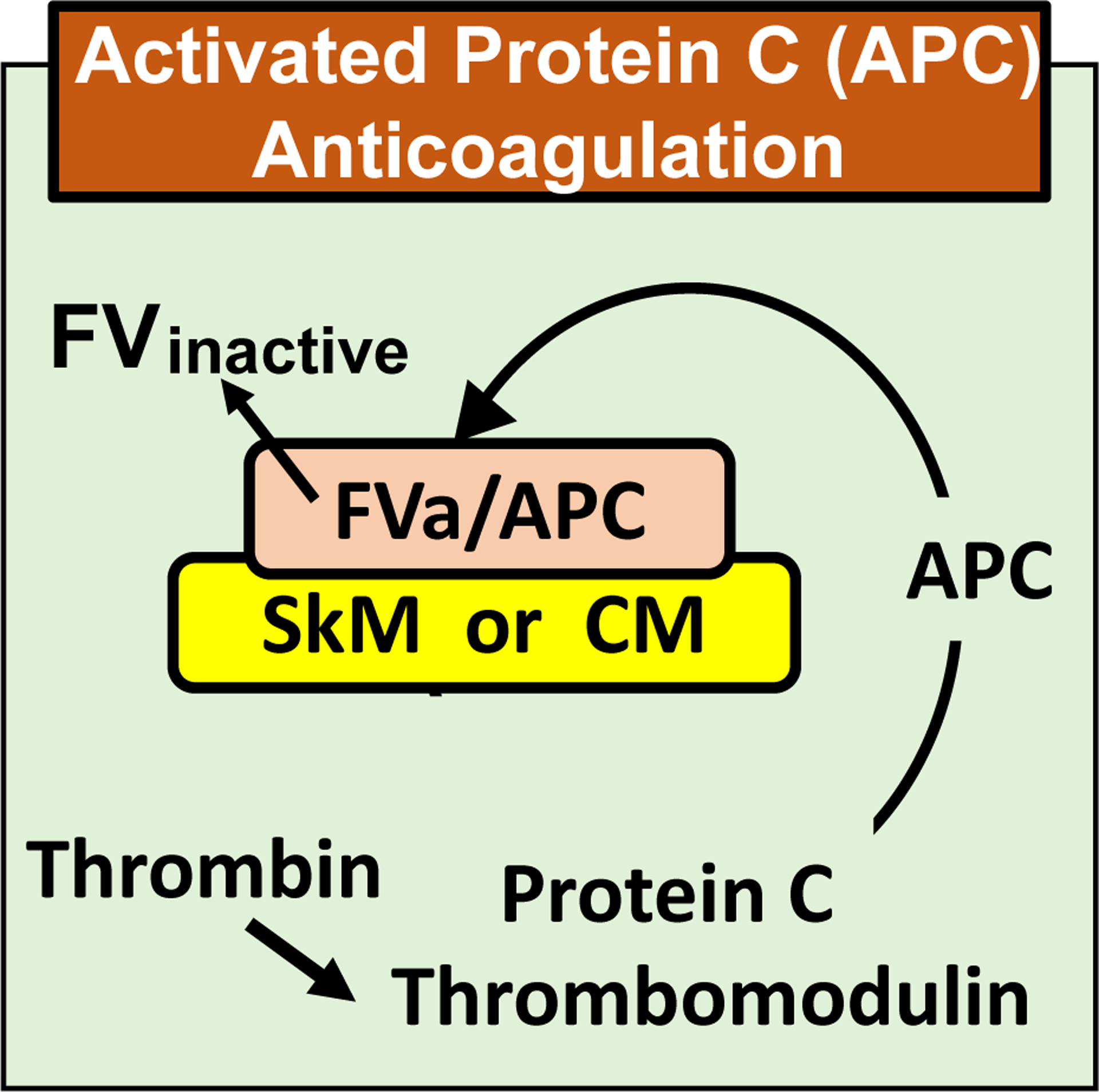
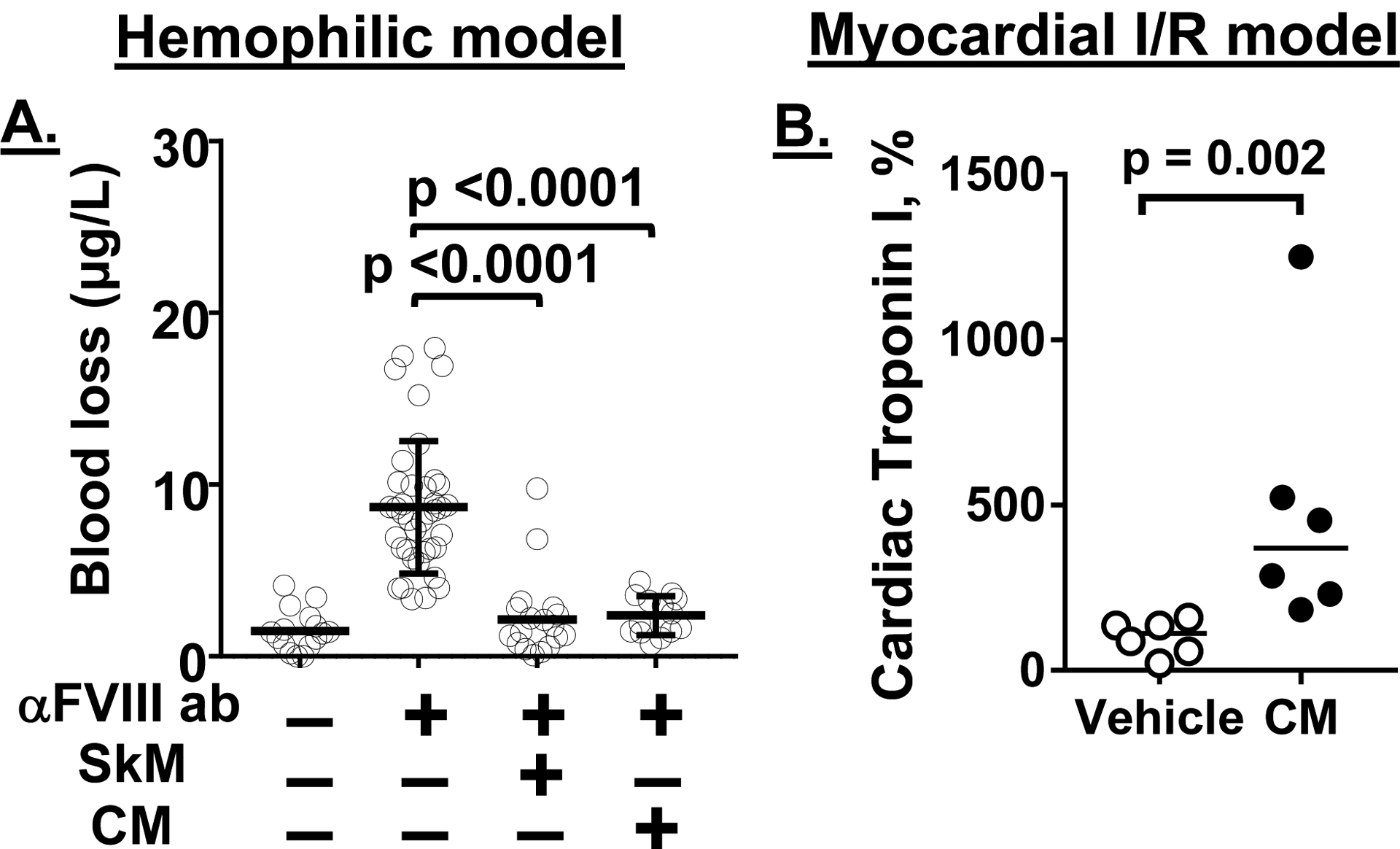
Essentials Striated muscle myosins can promote prothrombin activation by FXa or FVa inactivation by APC. Cardiac myosin and skeletal muscle myosin are pro-hemostatic in murine tail cut bleeding models. Infused cardiac myosin exacerbates myocardial injury caused by myocardial ischemia reperfusion. Skeletal muscle myosin isoforms that circulate in human plasma can be grouped into 3 phenotypes. ABSTRACT: Two striated muscle myosins, namely skeletal muscle myosin (SkM) and cardiac myosin (CM), may potentially contribute to physiologic mechanisms for regulation of thrombosis and hemostasis. Thrombin is generated from activation of prothrombin by the prothrombinase (IIase) complex comprising factor Xa, factor Va, and Ca++ ions located on surfaces where these factors are assembled. We discovered that SkM and CM, which are abundant motor proteins in skeletal and cardiac muscles, can provide a surface for thrombin generation by the prothrombinase complex without any apparent requirement for phosphatidylserine or lipids. These myosins can also provide a surface that supports the inactivation of factor Va by activated protein C/protein S, resulting in negative feedback downregulation of thrombin generation. Although the physiologic significance of these reactions remains to be established for humans, substantive insights may be gleaned from murine studies. In mice, exogenously infused SkM and CM can promote hemostasis as they are capable of reducing tail cut bleeding. In a murine myocardial ischemia-reperfusion injury model, exogenously infused CM exacerbates myocardial infarction damage. Studies of human plasmas show that SkM antigen isoforms of different MWs circulate in human plasma, and they can be used to identify three plasma SkM phenotypes. A pilot clinical study showed that one SkM isoform pattern appeared to be linked to isolated pulmonary embolism. These discoveries enable multiple preclinical and clinical studies of SkM and CM, which should provide novel mechanistic insights with potential translational relevance for the roles of CM and SkM in the pathobiology of hemostasis and thrombosis.
Keywords: blood coagulation; factor X; myocardial infarction; myosin; prothrombinase; thrombin.
© 2020 International Society on Thrombosis and Haemostasis.
Conflict of interest statement
CONFLICTS OF INTEREST
None of the authors has a conflict of interest to declare with regard to the content of this review.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
