

Viruses harness YxxØ motif to interact with host AP2M1 for replication: A vulnerable broad-spectrum antiviral target
- PMID: 32923629
- PMCID: PMC7455044
- DOI: 10.1126/sciadv.aba7910
Viruses harness YxxØ motif to interact with host AP2M1 for replication: A vulnerable broad-spectrum antiviral target
Abstract
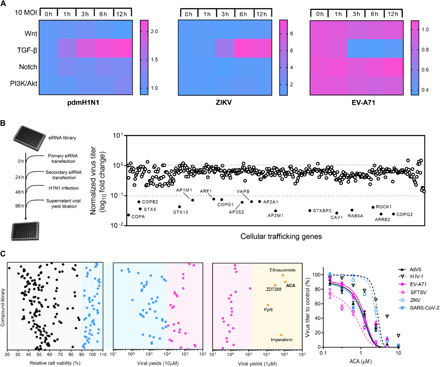
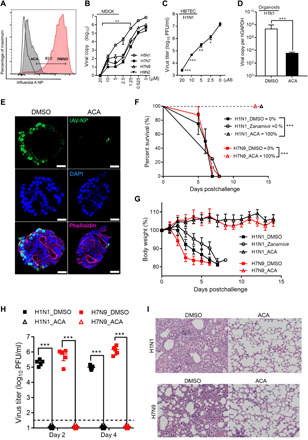
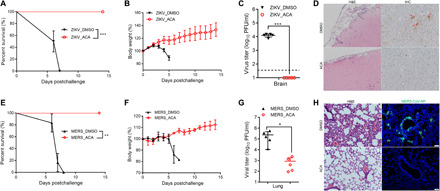
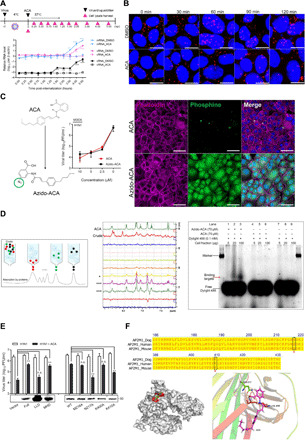
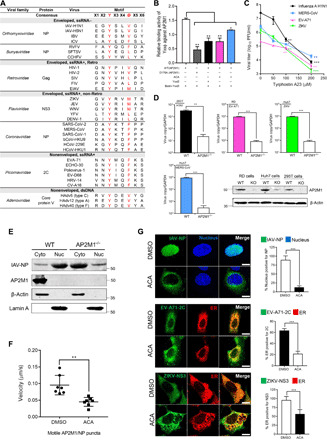
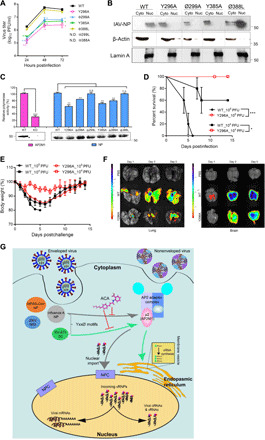
Targeting a universal host protein exploited by most viruses would be a game-changing strategy that offers broad-spectrum solution and rapid pandemic control including the current COVID-19. Here, we found a common YxxØ-motif of multiple viruses that exploits host AP2M1 for intracellular trafficking. A library chemical, N-(p-amylcinnamoyl)anthranilic acid (ACA), was identified to interrupt AP2M1-virus interaction and exhibit potent antiviral efficacy against a number of viruses in vitro and in vivo, including the influenza A viruses (IAVs), Zika virus (ZIKV), human immunodeficiency virus, and coronaviruses including MERS-CoV and SARS-CoV-2. YxxØ mutation, AP2M1 depletion, or disruption by ACA causes incorrect localization of viral proteins, which is exemplified by the failure of nuclear import of IAV nucleoprotein and diminished endoplasmic reticulum localization of ZIKV-NS3 and enterovirus-A71-2C proteins, thereby suppressing viral replication. Our study reveals an evolutionarily conserved mechanism of protein-protein interaction between host and virus that can serve as a broad-spectrum antiviral target.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
Similar articles
-
Ivermectin as a Broad-Spectrum Host-Directed Antiviral: The Real Deal?Cells. 2020 Sep 15;9(9):2100. doi: 10.3390/cells9092100. Cells. 2020. PMID: 32942671 Free PMC article. Review.
-
Novel and potent inhibitors targeting DHODH are broad-spectrum antivirals against RNA viruses including newly-emerged coronavirus SARS-CoV-2.Protein Cell. 2020 Oct;11(10):723-739. doi: 10.1007/s13238-020-00768-w. Epub 2020 Aug 4. Protein Cell. 2020. PMID: 32754890 Free PMC article.
-
A Carbohydrate-Binding Protein from the Edible Lablab Beans Effectively Blocks the Infections of Influenza Viruses and SARS-CoV-2.Cell Rep. 2020 Aug 11;32(6):108016. doi: 10.1016/j.celrep.2020.108016. Epub 2020 Jul 24. Cell Rep. 2020. PMID: 32755598 Free PMC article.
-
3C-like protease inhibitors block coronavirus replication in vitro and improve survival in MERS-CoV-infected mice.Sci Transl Med. 2020 Aug 19;12(557):eabc5332. doi: 10.1126/scitranslmed.abc5332. Epub 2020 Aug 3. Sci Transl Med. 2020. PMID: 32747425 Free PMC article.
-
Broad Spectrum Antiviral Agent Niclosamide and Its Therapeutic Potential.ACS Infect Dis. 2020 May 8;6(5):909-915. doi: 10.1021/acsinfecdis.0c00052. Epub 2020 Mar 10. ACS Infect Dis. 2020. PMID: 32125140 Free PMC article. Review.
Cited by
-
D3AI-CoV: a deep learning platform for predicting drug targets and for virtual screening against COVID-19.Brief Bioinform. 2022 May 13;23(3):bbac147. doi: 10.1093/bib/bbac147. Brief Bioinform. 2022. PMID: 35443040 Free PMC article.
-
SaeR as a novel target for antivirulence therapy against Staphylococcus aureus.Emerg Microbes Infect. 2023 Dec;12(2):2254415. doi: 10.1080/22221751.2023.2254415. Emerg Microbes Infect. 2023. PMID: 37671453 Free PMC article.
-
Development of Three-Dimensional Human Intestinal Organoids as a Physiologically Relevant Model for Characterizing the Viral Replication Kinetics and Antiviral Susceptibility of Enteroviruses.Biomedicines. 2021 Jan 18;9(1):88. doi: 10.3390/biomedicines9010088. Biomedicines. 2021. PMID: 33477611 Free PMC article.
-
Large scale discovery of coronavirus-host factor protein interaction motifs reveals SARS-CoV-2 specific mechanisms and vulnerabilities.Nat Commun. 2021 Nov 19;12(1):6761. doi: 10.1038/s41467-021-26498-z. Nat Commun. 2021. PMID: 34799561 Free PMC article.
-
Identification of the SARS-unique domain of SARS-CoV-2 as an antiviral target.Nat Commun. 2023 Jul 6;14(1):3999. doi: 10.1038/s41467-023-39709-6. Nat Commun. 2023. PMID: 37414753 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
