

Honeybee venom and melittin suppress growth factor receptor activation in HER2-enriched and triple-negative breast cancer
- PMID: 32923684
- PMCID: PMC7463160
- DOI: 10.1038/s41698-020-00129-0
Honeybee venom and melittin suppress growth factor receptor activation in HER2-enriched and triple-negative breast cancer
Abstract
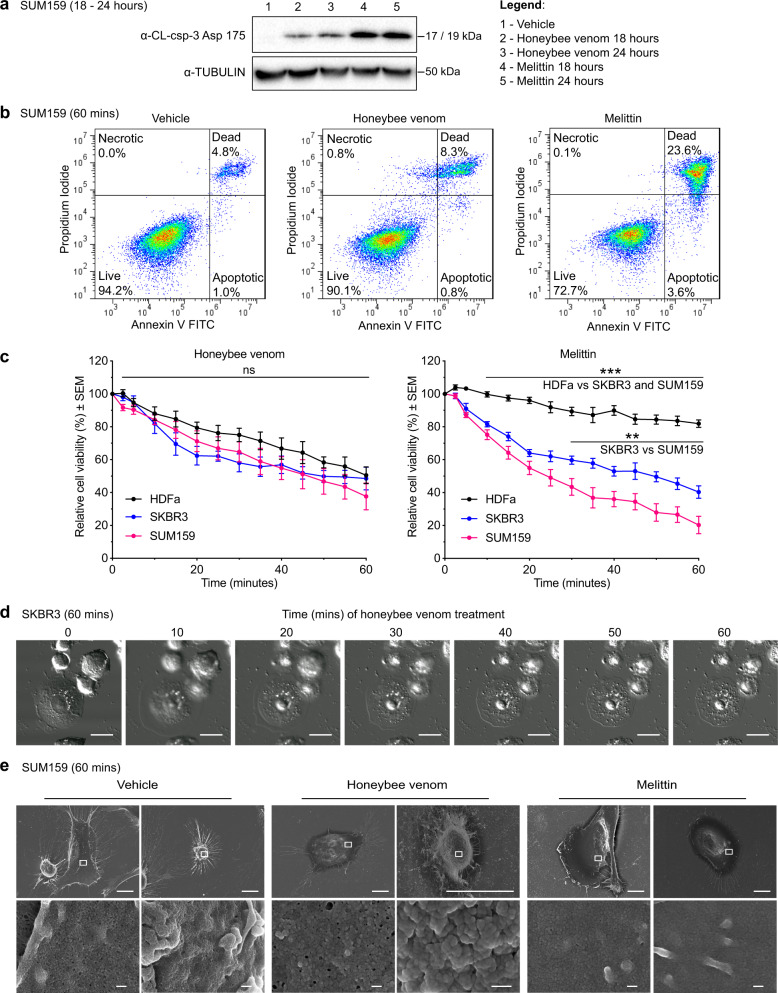
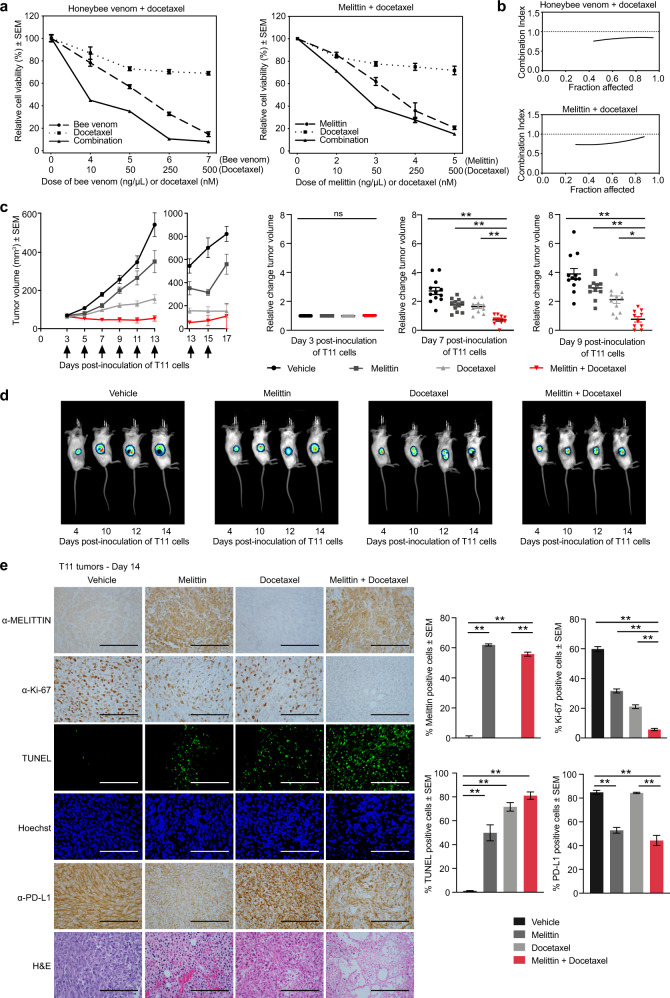
Despite decades of study, the molecular mechanisms and selectivity of the biomolecular components of honeybee (Apis mellifera) venom as anticancer agents remain largely unknown. Here, we demonstrate that honeybee venom and its major component melittin potently induce cell death, particularly in the aggressive triple-negative and HER2-enriched breast cancer subtypes. Honeybee venom and melittin suppress the activation of EGFR and HER2 by interfering with the phosphorylation of these receptors in the plasma membrane of breast carcinoma cells. Mutational studies reveal that a positively charged C-terminal melittin sequence mediates plasma membrane interaction and anticancer activity. Engineering of an RGD motif further enhances targeting of melittin to malignant cells with minimal toxicity to normal cells. Lastly, administration of melittin enhances the effect of docetaxel in suppressing breast tumor growth in an allograft model. Our work unveils a molecular mechanism underpinning the anticancer selectivity of melittin, and outlines treatment strategies to target aggressive breast cancers.
Keywords: Breast cancer; Molecular medicine.
© The Author(s) 2020.
Conflict of interest statement
Competing interestsK.P. received funding from Promega, BMG Labtech, and Dimerix as ARC Linkage Grant participating organizations. These participating organizations played no role in any aspect of the conception or design of the research, collection, analysis and interpretation of the results, or writing and editing of the paper. K.P. is the chief scientific advisor of Dimerix, of which he maintains a shareholding. Remaining authors declare no competing interests.
Figures



References
-
- Son DJ, et al. Therapeutic application of anti-arthritis, pain-releasing, and anti-cancer effects of bee venom and its constituent compounds. Pharmacol. Ther. 2007;115:246–270. - PubMed
-
- Park S-C, et al. Investigation of toroidal pore and oligomerization by melittin using transmission electron microscopy. Biochem. Biophys. Res. Commun. 2006;343:222–228. - PubMed
-
- Lyu Y, Zhu X, Xiang N, Narsimhan G. Molecular Dynamics Study of Pore Formation by Melittin in a 1,2-Dioleoyl- sn -glycero-3-phosphocholine and 1,2-Di(9 Z -octadecenoyl)- sn -glycero-3-phospho-(1′- rac -glycerol) Mixed Lipid Bilayer. Ind. Eng. Chem. Res. 2015;54:10275–10283.
-
- Terwilliger TC, Eisenberg D. The structure of melittin. II. Interpretation of the structure. J. Biol. Chem. 1982;257:6016–6022. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
