

An Ixodes scapularis Protein Disulfide Isomerase Contributes to Borrelia burgdorferi Colonization of the Vector
- PMID: 32928964
- PMCID: PMC7671890
- DOI: 10.1128/IAI.00426-20
An Ixodes scapularis Protein Disulfide Isomerase Contributes to Borrelia burgdorferi Colonization of the Vector
Abstract
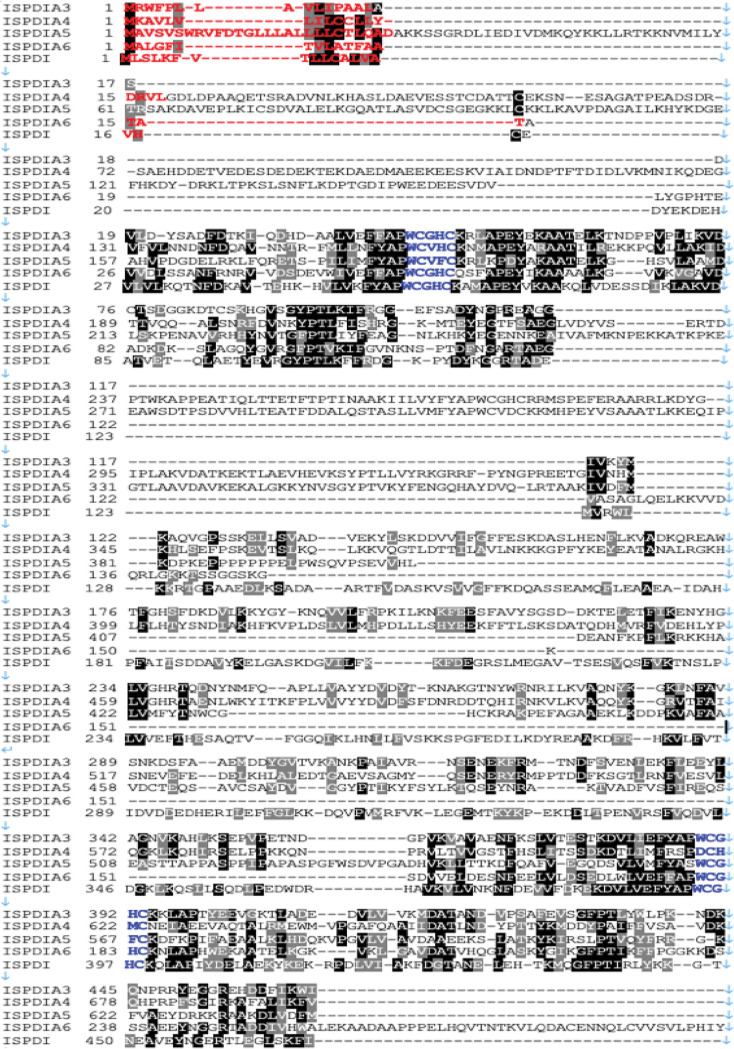
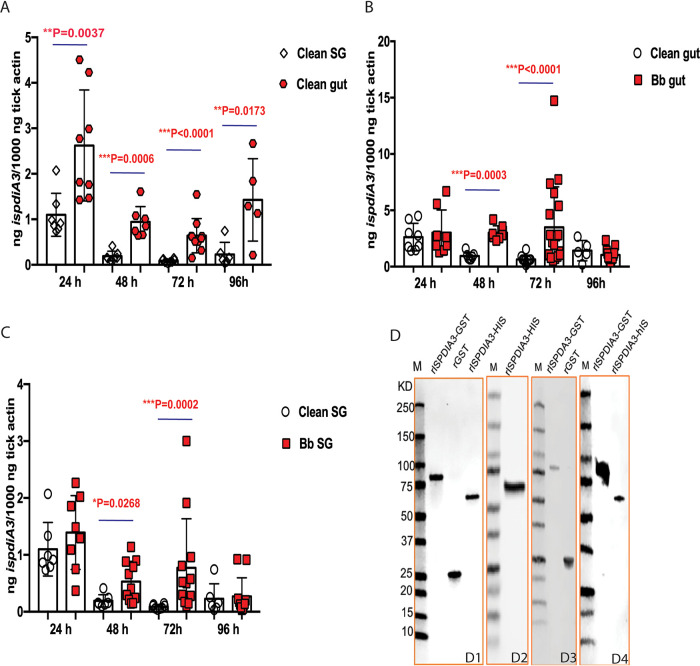
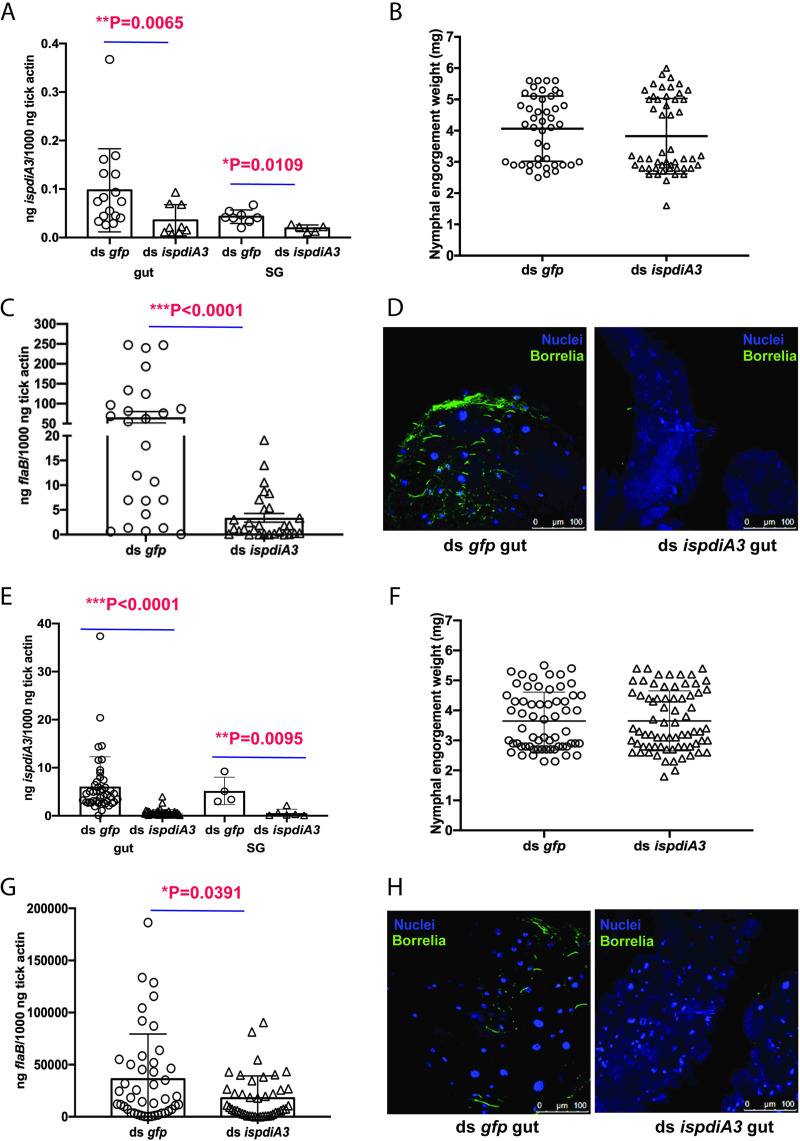
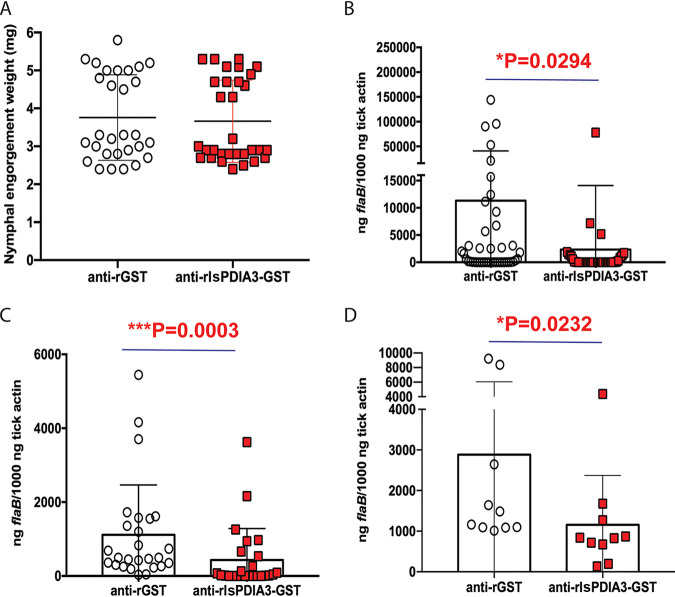
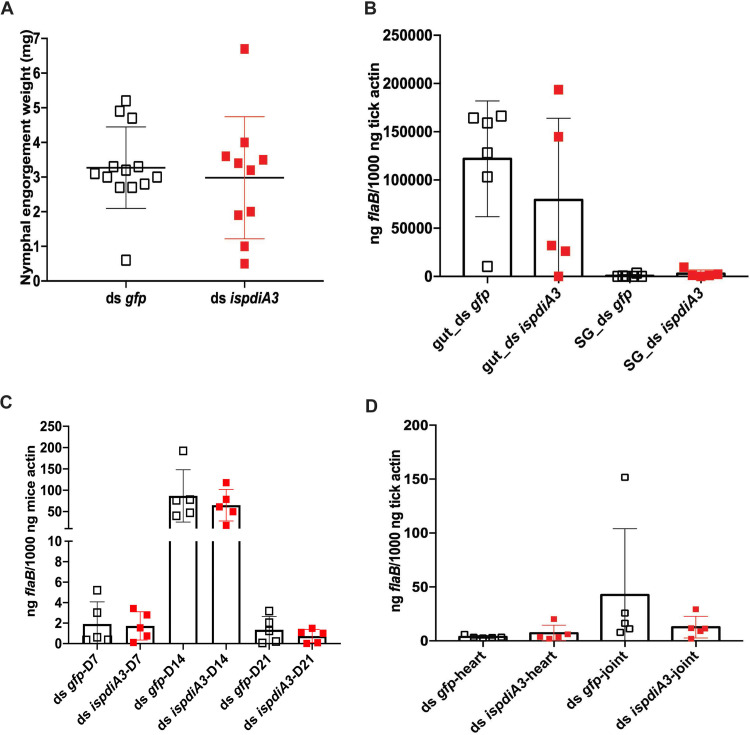
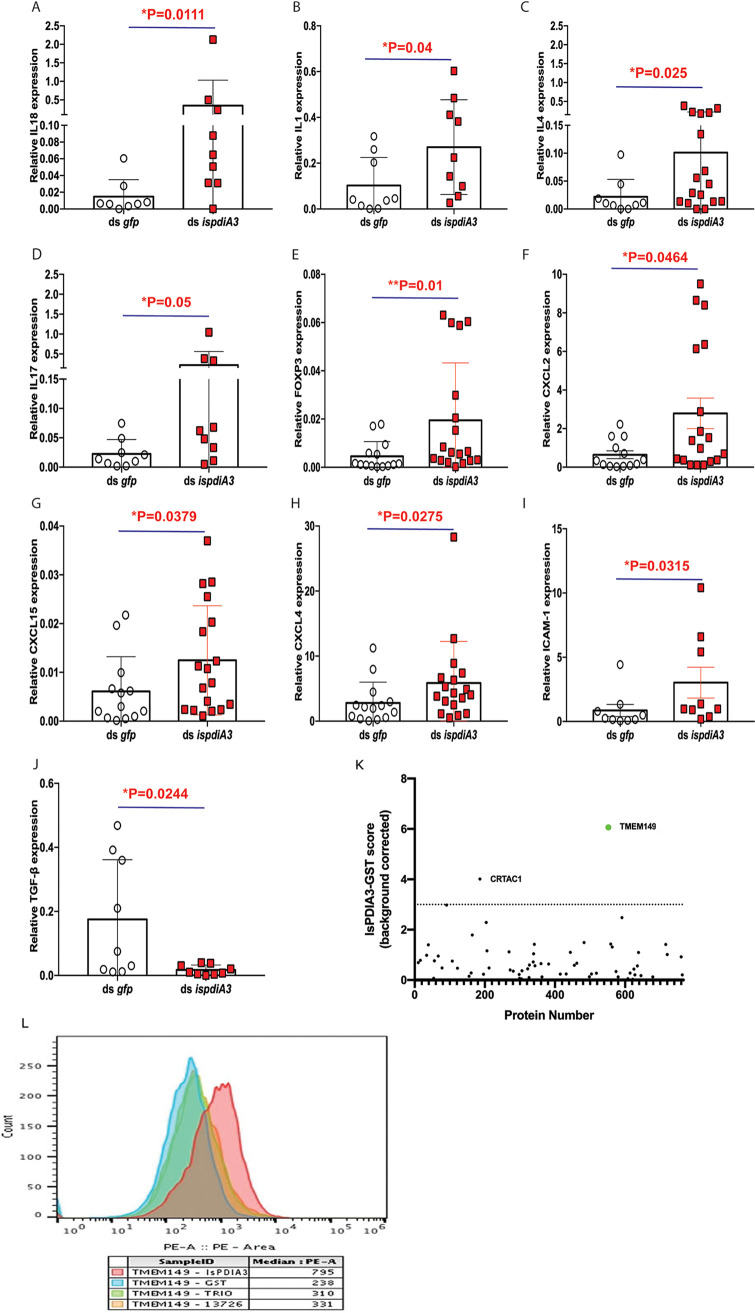
Borrelia burgdorferi causes Lyme disease, the most common tick-transmitted illness in North America. When Ixodes scapularis feed on an infected vertebrate host, spirochetes enter the tick gut along with the bloodmeal and colonize the vector. Here, we show that a secreted tick protein,
Keywords: Borrelia burgdorferi; Ixodes scapularis; colonization; protein disulfide isomerase.
Copyright © 2020 Cao et al.
Figures


References
-
- Hair JA, Bowman J, Sauer J. 1986. Morphology, physiology and behavioural biology of ticks. Ellis Horwood, Chichester, UK.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
