

Unraveling the sugar code: the role of microbial extracellular glycans in plant-microbe interactions
- PMID: 32929496
- PMCID: PMC7816849
- DOI: 10.1093/jxb/eraa414
Unraveling the sugar code: the role of microbial extracellular glycans in plant-microbe interactions
Abstract
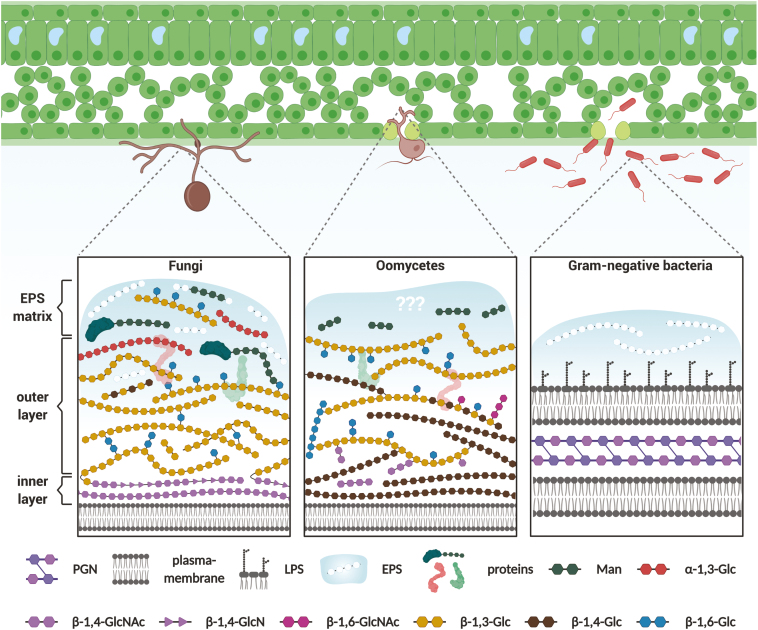
To defend against microbial invaders but also to establish symbiotic programs, plants need to detect the presence of microbes through the perception of molecular signatures characteristic of a whole class of microbes. Among these molecular signatures, extracellular glycans represent a structurally complex and diverse group of biomolecules that has a pivotal role in the molecular dialog between plants and microbes. Secreted glycans and glycoconjugates such as symbiotic lipochitooligosaccharides or immunosuppressive cyclic β-glucans act as microbial messengers that prepare the ground for host colonization. On the other hand, microbial cell surface glycans are important indicators of microbial presence. They are conserved structures normally exposed and thus accessible for plant hydrolytic enzymes and cell surface receptor proteins. While the immunogenic potential of bacterial cell surface glycoconjugates such as lipopolysaccharides and peptidoglycan has been intensively studied in the past years, perception of cell surface glycans from filamentous microbes such as fungi or oomycetes is still largely unexplored. To date, only few studies have focused on the role of fungal-derived cell surface glycans other than chitin, highlighting a knowledge gap that needs to be addressed. The objective of this review is to give an overview on the biological functions and perception of microbial extracellular glycans, primarily focusing on their recognition and their contribution to plant-microbe interactions.
Keywords: Cell wall; chitin; extracellular polysaccharides; glucan; immunity; matrix; microbes; symbiosis.
© The Author(s) 2020. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Acosta-Jurado S, Rodriguez-Navarro DN, Kawaharada Y, et al. 2016. Sinorhizobium fredii HH103 invades Lotus burttii by crack entry in a nod factor- and surface polysaccharide-dependent manner. Molecular Plant-Microbe Interactions 29, 925–937. - PubMed
-
- Amor BB, Shaw SL, Oldroyd GE, Maillet F, Penmetsa RV, Cook D, Long SR, Denarie J, Gough C. 2003. The NFP locus of Medicago truncatula controls an early step of Nod factor signal transduction upstream of a rapid calcium flux and root hair deformation. The Plant Journal 34, 495–506. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
