

Looking back and looking forward: contributions of electron microscopy to the structural cell biology of gametes and fertilization
- PMID: 32931719
- PMCID: PMC7536082
- DOI: 10.1098/rsob.200186
Looking back and looking forward: contributions of electron microscopy to the structural cell biology of gametes and fertilization
Abstract
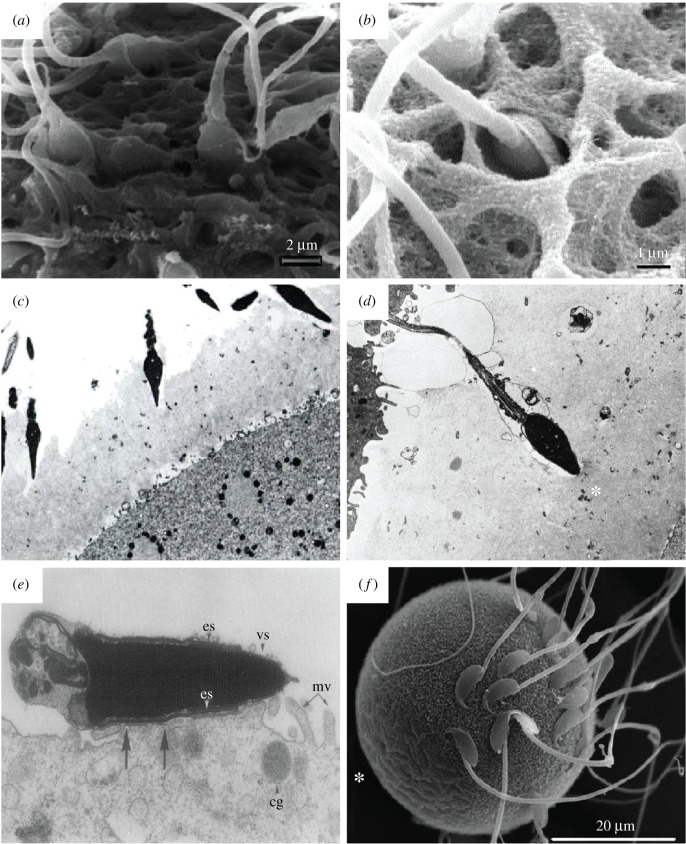
Mammalian gametes-the sperm and the egg-represent opposite extremes of cellular organization and scale. Studying the ultrastructure of gametes is crucial to understanding their interactions, and how to manipulate them in order to either encourage or prevent their union. Here, we survey the prominent electron microscopy (EM) techniques, with an emphasis on considerations for applying them to study mammalian gametes. We review how conventional EM has provided significant insight into gamete ultrastructure, but also how the harsh sample preparation methods required preclude understanding at a truly molecular level. We present recent advancements in cryo-electron tomography that provide an opportunity to image cells in a near-native state and at unprecedented levels of detail. New and emerging cellular EM techniques are poised to rekindle exploration of fundamental questions in mammalian reproduction, especially phenomena that involve complex membrane remodelling and protein reorganization. These methods will also allow novel lines of enquiry into problems of practical significance, such as investigating unexplained causes of human infertility and improving assisted reproductive technologies for biodiversity conservation.
Keywords: cryo-electron tomography; cryo-focused ion beam milling; egg; electron microscopy; fertilization; sperm.
Conflict of interest statement
We declare we have no competing interests.
Figures





References
-
- van Leeuwenhoek A. 1677. Observationes D. Anthonii Lewenhoeck, de natis'e semine genitali animalculis. Phil. Trans. R. Soc. 12, 1040–1046. ( 10.1098/rstl.1677.0068) - DOI
-
- von Baer KE. 1827. De ovi mammalium et hominis genesi (On the genesis of the ovum of mammals and of man). Leipzig, Germany: Leopold Voss.
-
- Hertwig O. 1876. Beiträge zur kenntniss der bildung, befruchtung und theilung des thierischen eies. Morph. Jahrb. 1, 347–434.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources