

Sensory Experience Engages Microglia to Shape Neural Connectivity through a Non-Phagocytic Mechanism
- PMID: 32931754
- PMCID: PMC7666095
- DOI: 10.1016/j.neuron.2020.08.002
Sensory Experience Engages Microglia to Shape Neural Connectivity through a Non-Phagocytic Mechanism
Abstract
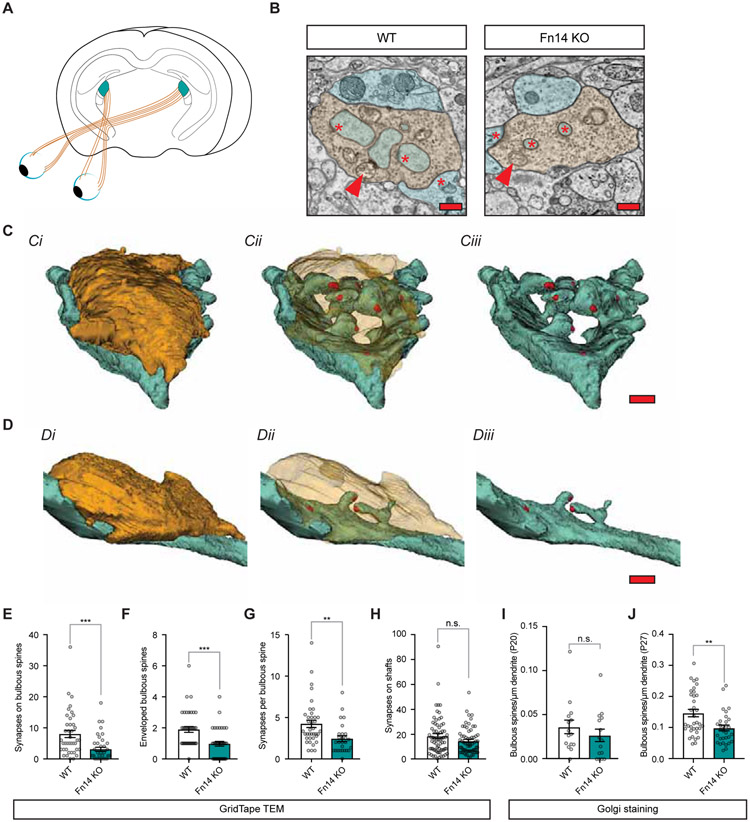
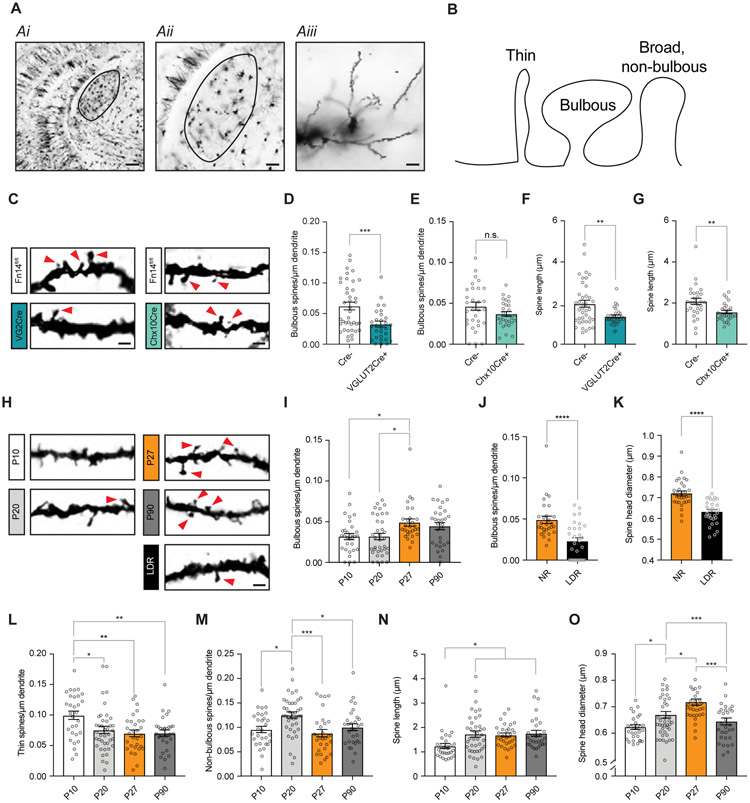
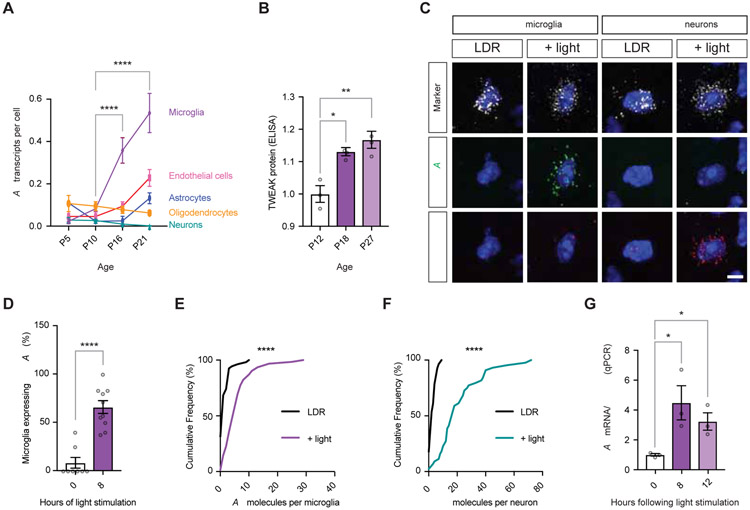
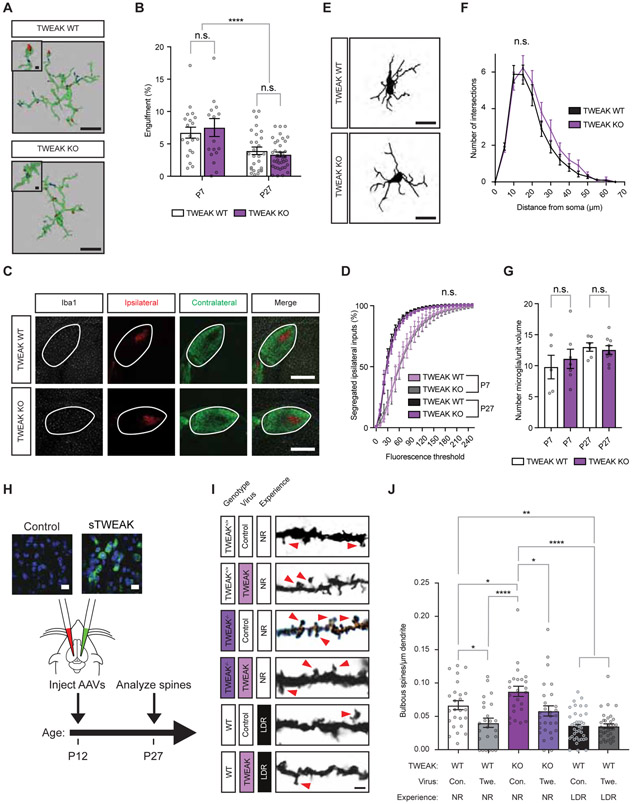
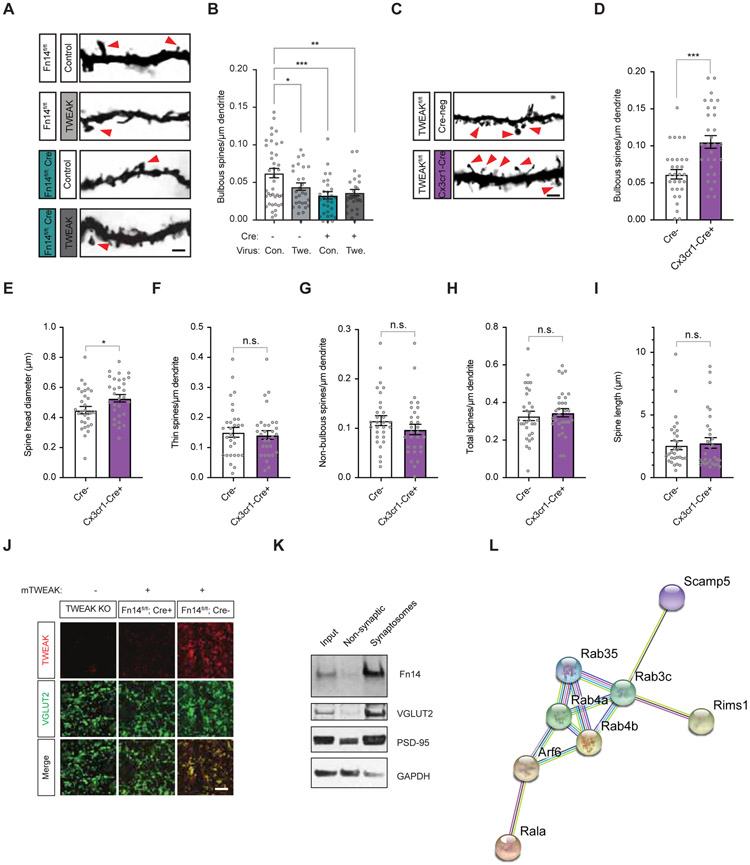
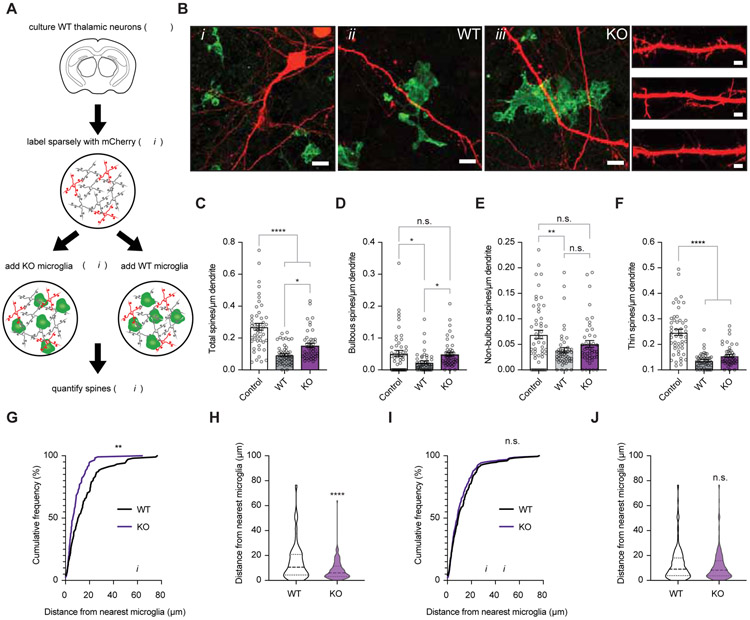
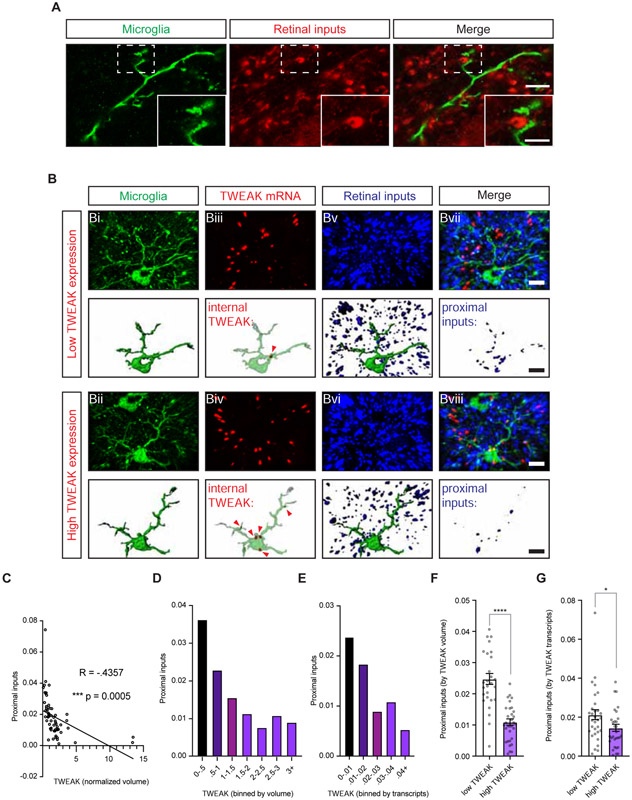
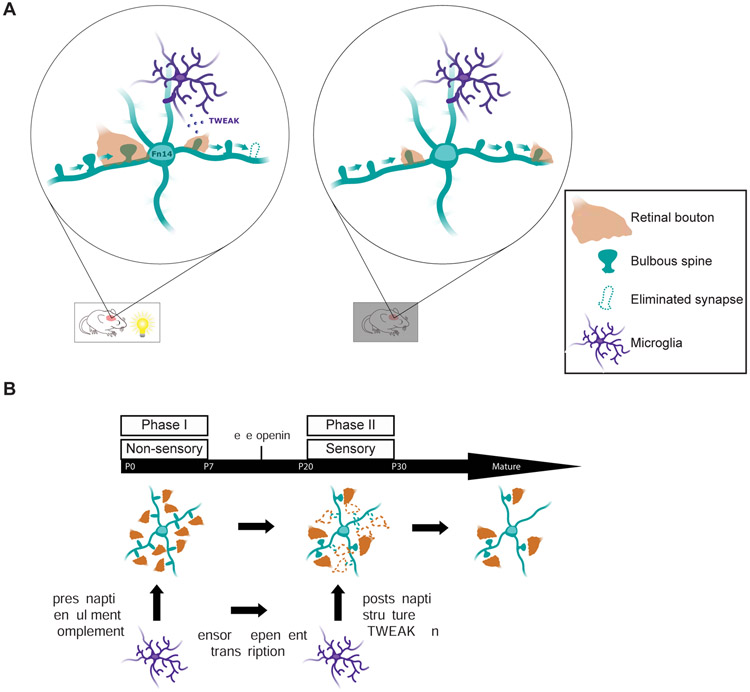
Sensory experience remodels neural circuits in the early postnatal brain through mechanisms that remain to be elucidated. Applying a new method of ultrastructural analysis to the retinogeniculate circuit, we find that visual experience alters the number and structure of synapses between the retina and the thalamus. These changes require vision-dependent transcription of the receptor Fn14 in thalamic relay neurons and the induction of its ligand TWEAK in microglia. Fn14 functions to increase the number of bulbous spine-associated synapses at retinogeniculate connections, likely contributing to the strengthening of the circuit that occurs in response to visual experience. However, at retinogeniculate connections near TWEAK-expressing microglia, TWEAK signals via Fn14 to restrict the number of bulbous spines on relay neurons, leading to the elimination of a subset of connections. Thus, TWEAK and Fn14 represent an intercellular signaling axis through which microglia shape retinogeniculate connectivity in response to sensory experience.
Keywords: dendritic spine; development; microglia; pruning; sensory experience; synapse; synaptic refinement.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests L.C.B. and K.A.E. are employees and shareholders of Biogen.
Figures
Comment in
-
Tweaking synapses.Nat Rev Neurosci. 2020 Nov;21(11):594. doi: 10.1038/s41583-020-00389-6. Nat Rev Neurosci. 2020. PMID: 32989276 No abstract available.
-
Microglia Tweak Retinogeniculate Pathways during Visual Circuit Refinement.Neuron. 2020 Nov 11;108(3):397-399. doi: 10.1016/j.neuron.2020.10.018. Neuron. 2020. PMID: 33181071
References
-
- Cardona AE, Huang D, Sasse ME, and Ransohoff RM (2006). Isolation of murine microglial cells for RNA analysis or flow cytometry. Nat Protoc 1, 1947–1951. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
