

A Genome-Scale Metabolic Model of Thalassiosira pseudonana CCMP 1335 for a Systems-Level Understanding of Its Metabolism and Biotechnological Potential
- PMID: 32932853
- PMCID: PMC7563145
- DOI: 10.3390/microorganisms8091396
A Genome-Scale Metabolic Model of Thalassiosira pseudonana CCMP 1335 for a Systems-Level Understanding of Its Metabolism and Biotechnological Potential
Abstract
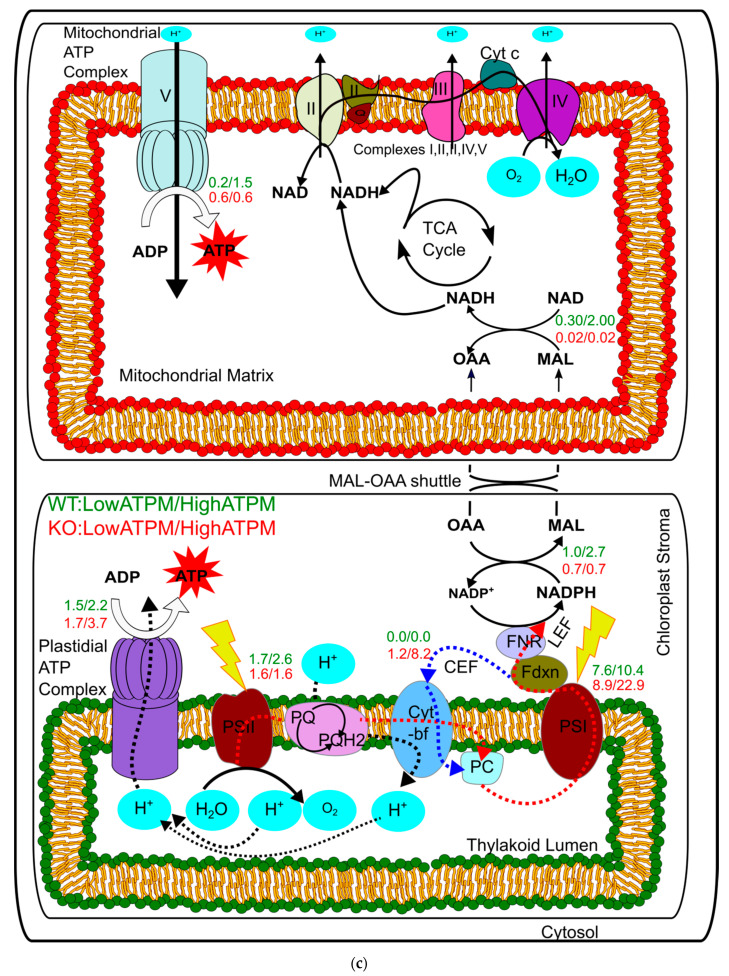
Thalassiosira pseudonana is a transformable and biotechnologically promising model diatom with an ability to synthesise nutraceuticals such as fucoxanthin and store a significant amount of polyglucans and lipids including omega-3 fatty acids. While it was the first diatom to be sequenced, a systems-level analysis of its metabolism has not been done yet. This work presents first comprehensive, compartmentalized, and functional genome-scale metabolic model of the marine diatom Thalassiosira pseudonana CCMP 1335, which we have termed iThaps987. The model includes 987 genes, 2477 reactions, and 2456 metabolites. Comparison with the model of another diatom Phaeodactylum tricornutum revealed presence of 183 unique enzymes (belonging primarily to amino acid, carbohydrate, and lipid metabolism) in iThaps987. Model simulations showed a typical C3-type photosynthetic carbon fixation and suggested a preference of violaxanthin-diadinoxanthin pathway over violaxanthin-neoxanthin pathway for the production of fucoxanthin. Linear electron flow was found be active and cyclic electron flow was inactive under normal phototrophic conditions (unlike green algae and plants), validating the model predictions with previous reports. Investigation of the model for the potential of Thalassiosira pseudonana CCMP 1335 to produce other industrially useful compounds suggest iso-butanol as a foreign compound that can be synthesized by a single-gene addition. This work provides novel insights about the metabolism and potential of the organism and will be helpful to further investigate its metabolism and devise metabolic engineering strategies for the production of various compounds.
Keywords: diatom; flux analysis; fucoxanthin; heterologous products; photosynthesis.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection; analyses; or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures






References
-
- Nelson D.M., Tréguer P., Brzezinski M.A., Leynaert A., Quéguiner B. Production and dissolution of biogenic silica in the ocean: Revised global estimates, comparison with regional data and relationship to biogenic sedimentation. Glob. Biogeochem. Cycles. 1995;9:359–372. doi: 10.1029/95GB01070. - DOI
-
- Hildebrand M., Davis A.K., Smith S.R., Traller J.C., Abbriano R. The place of diatoms in the biofuels industry. Biofuels. 2012;3:221–240. doi: 10.4155/bfs.11.157. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
