

MARCH8 Inhibits Ebola Virus Glycoprotein, Human Immunodeficiency Virus Type 1 Envelope Glycoprotein, and Avian Influenza Virus H5N1 Hemagglutinin Maturation
- PMID: 32934085
- PMCID: PMC7492737
- DOI: 10.1128/mBio.01882-20
MARCH8 Inhibits Ebola Virus Glycoprotein, Human Immunodeficiency Virus Type 1 Envelope Glycoprotein, and Avian Influenza Virus H5N1 Hemagglutinin Maturation
Abstract
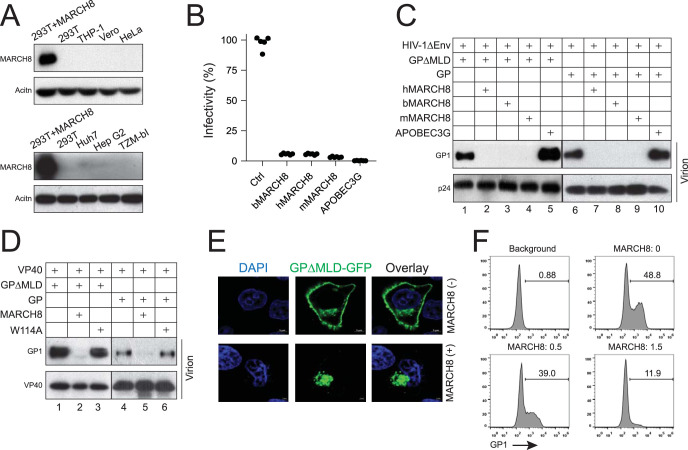
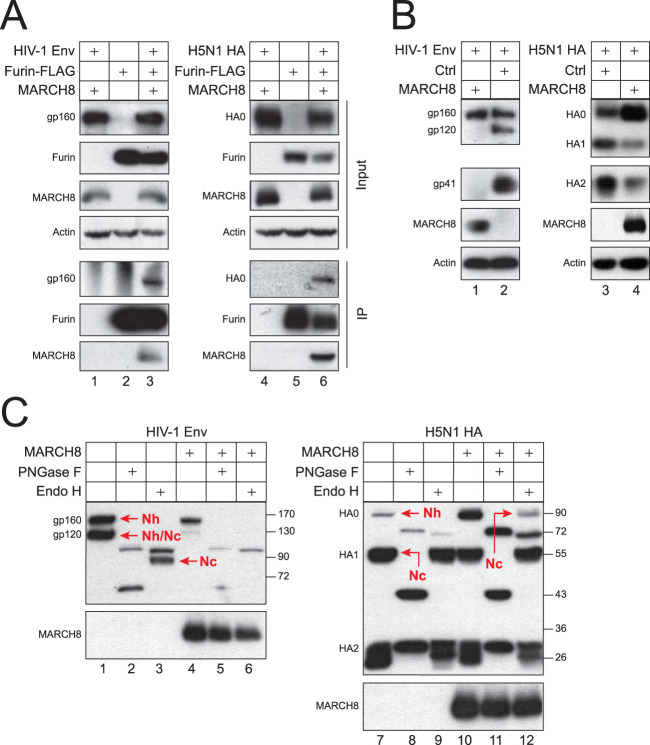
Membrane-associated RING-CH-type 8 (MARCH8) strongly blocks human immunodeficiency virus type 1 (HIV-1) envelope glycoprotein (Env) incorporation into virions by downregulating its cell surface expression, but the mechanism is still unclear. We now report that MARCH8 also blocks the Ebola virus (EBOV) glycoprotein (GP) incorporation via surface downregulation. To understand how these viral fusion proteins are downregulated, we investigated the effects of MARCH8 on EBOV GP maturation and externalization via the conventional secretion pathway. MARCH8 interacted with EBOV GP and furin when detected by immunoprecipitation and retained the GP/furin complex in the Golgi when their location was tracked by a bimolecular fluorescence complementation (BiFC) assay. MARCH8 did not reduce the GP expression or affect the GP modification by high-mannose N-glycans in the endoplasmic reticulum (ER), but it inhibited the formation of complex N-glycans on the GP in the Golgi. Additionally, the GP O-glycosylation and furin-mediated proteolytic cleavage were also inhibited. Moreover, we identified a novel furin cleavage site on EBOV GP and found that only those fully glycosylated GPs were processed by furin and incorporated into virions. Furthermore, the GP shedding and secretion were all blocked by MARCH8. MARCH8 also blocked the furin-mediated cleavage of HIV-1 Env (gp160) and the highly pathogenic avian influenza virus H5N1 hemagglutinin (HA). We conclude that MARCH8 has a very broad antiviral activity by prohibiting different viral fusion proteins from glycosylation and proteolytic cleavage in the Golgi, which inhibits their transport from the Golgi to the plasma membrane and incorporation into virions.IMPORTANCE Enveloped viruses express three classes of fusion proteins that are required for their entry into host cells via mediating virus and cell membrane fusion. Class I fusion proteins are produced from influenza viruses, retroviruses, Ebola viruses, and coronaviruses. They are first synthesized as a type I transmembrane polypeptide precursor that is subsequently glycosylated and oligomerized. Most of these precursors are cleaved en route to the plasma membrane by a cellular protease furin in the late secretory pathway, generating the trimeric N-terminal receptor-binding and C-terminal fusion subunits. Here, we show that a cellular protein, MARCH8, specifically inhibits the furin-mediated cleavage of EBOV GP, HIV-1 Env, and H5N1 HA. Further analyses uncovered that MARCH8 blocked the EBOV GP glycosylation in the Golgi and inhibited its transport from the Golgi to the plasma membrane. Thus, MARCH8 has a very broad antiviral activity by specifically inactivating different viral fusion proteins.
Keywords: Ebola; HIV; MARCH8; class I fusion protein; furin; glycosylation; influenza; viral envelope.
Copyright © 2020 Yu et al.
Figures






Similar articles
-
Human MARCH1, 2, and 8 block Ebola virus envelope glycoprotein cleavage via targeting furin P domain.J Med Virol. 2024 Feb;96(2):e29445. doi: 10.1002/jmv.29445. J Med Virol. 2024. PMID: 38299743
-
Mechanism of Viral Glycoprotein Targeting by Membrane-Associated RING-CH Proteins.mBio. 2021 Mar 16;12(2):e00219-21. doi: 10.1128/mBio.00219-21. mBio. 2021. PMID: 33727347 Free PMC article.
-
A Virion-Based Assay for Glycoprotein Thermostability Reveals Key Determinants of Filovirus Entry and Its Inhibition.J Virol. 2020 Aug 31;94(18):e00336-20. doi: 10.1128/JVI.00336-20. Print 2020 Aug 31. J Virol. 2020. PMID: 32611759 Free PMC article.
-
Maturation of HIV envelope glycoprotein precursors by cellular endoproteases.Biochim Biophys Acta. 2000 Nov 10;1469(3):121-32. doi: 10.1016/s0304-4157(00)00014-9. Biochim Biophys Acta. 2000. PMID: 11063880 Review.
-
MARCH8: the tie that binds to viruses.FEBS J. 2022 Jul;289(13):3642-3654. doi: 10.1111/febs.16017. Epub 2021 May 31. FEBS J. 2022. PMID: 33993615 Review.
Cited by
-
Antiviral mechanisms of guanylate-binding protein 5: versatile inhibition of multiple viral glycoproteins.mBio. 2024 Nov 13;15(11):e0237424. doi: 10.1128/mbio.02374-24. Epub 2024 Oct 15. mBio. 2024. PMID: 39404361 Free PMC article.
-
HSC70 Inhibits Spring Viremia of Carp Virus Replication by Inducing MARCH8-Mediated Lysosomal Degradation of G Protein.Front Immunol. 2021 Sep 29;12:724403. doi: 10.3389/fimmu.2021.724403. eCollection 2021. Front Immunol. 2021. PMID: 34659210 Free PMC article.
-
MARCH8 Restricts Influenza A Virus Infectivity but Does Not Downregulate Viral Glycoprotein Expression at the Surface of Infected Cells.mBio. 2021 Oct 26;12(5):e0148421. doi: 10.1128/mBio.01484-21. Epub 2021 Sep 14. mBio. 2021. PMID: 34517760 Free PMC article.
-
Role of Host-Mediated Post-Translational Modifications (PTMs) in RNA Virus Pathogenesis.Int J Mol Sci. 2020 Dec 30;22(1):323. doi: 10.3390/ijms22010323. Int J Mol Sci. 2020. PMID: 33396899 Free PMC article. Review.
-
Deglycosylation of SLAMF7 in breast cancers enhances phagocytosis.Am J Cancer Res. 2022 Oct 15;12(10):4721-4736. eCollection 2022. Am J Cancer Res. 2022. PMID: 36381324 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
