

Characterization and mitigation of gene expression burden in mammalian cells
- PMID: 32934213
- PMCID: PMC7492461
- DOI: 10.1038/s41467-020-18392-x
Characterization and mitigation of gene expression burden in mammalian cells
Abstract
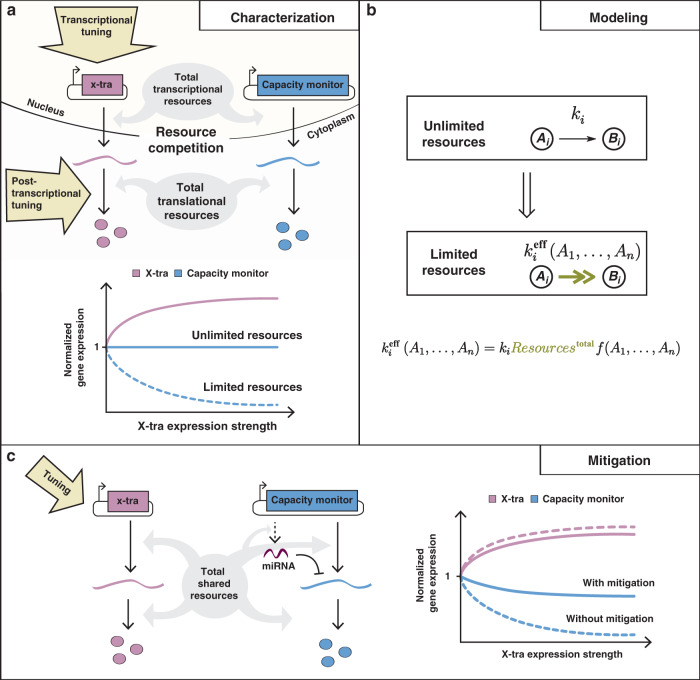
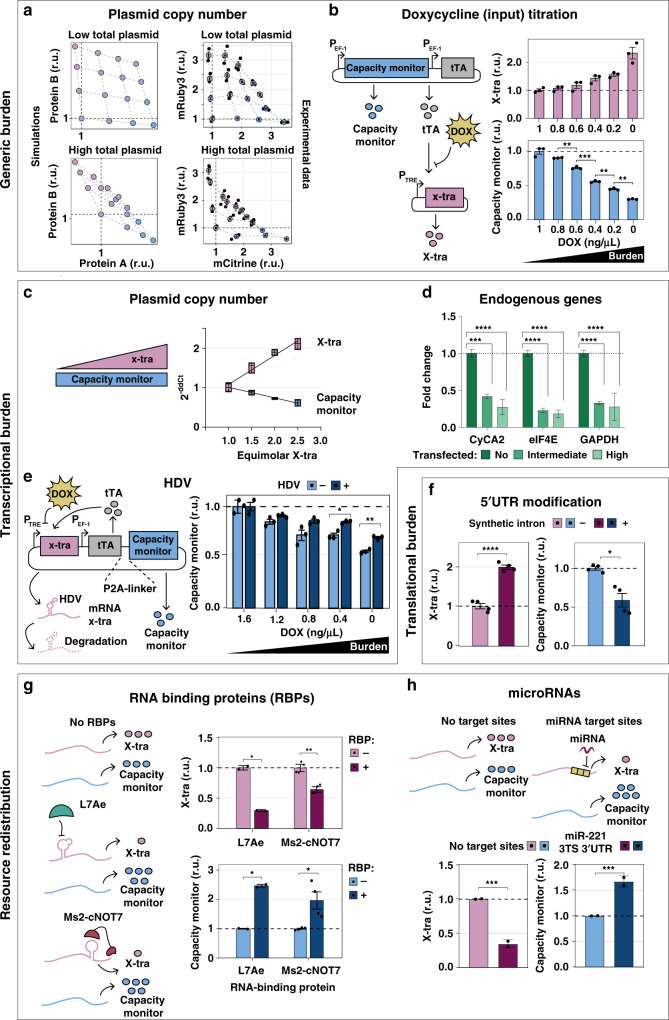
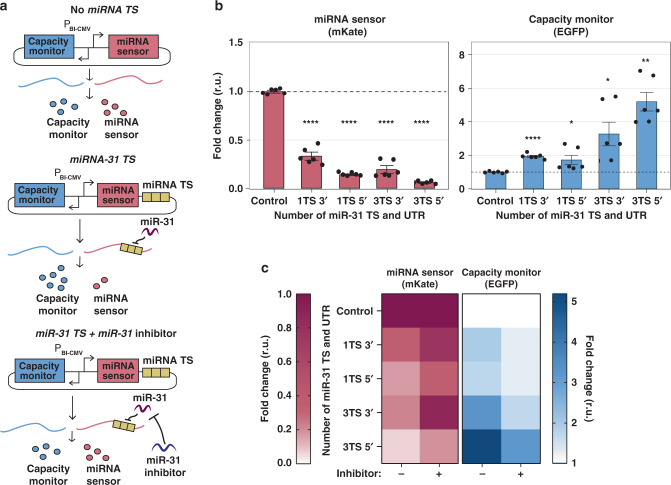
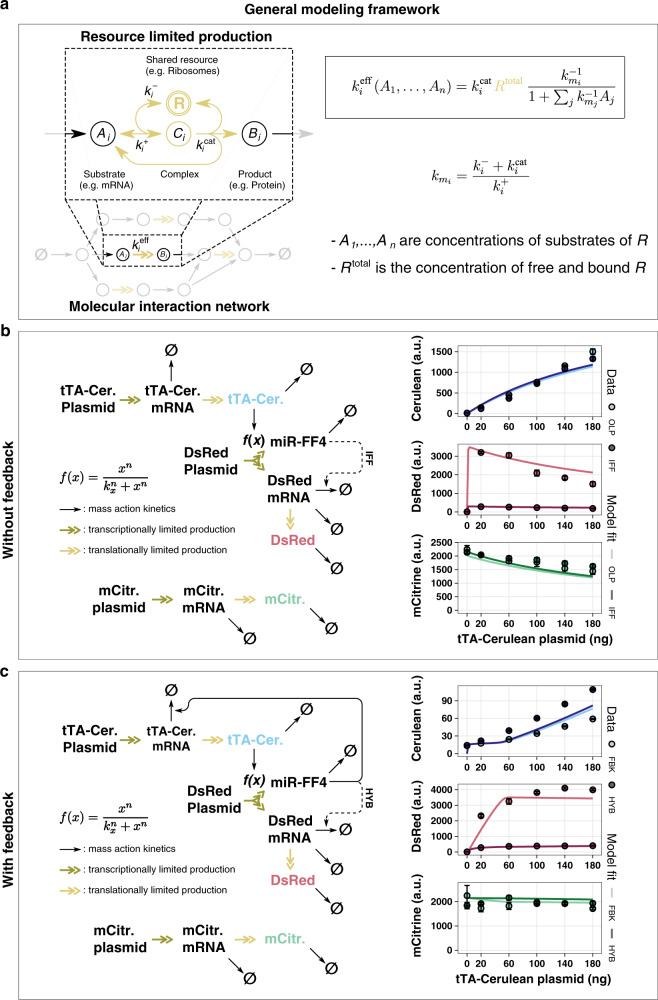
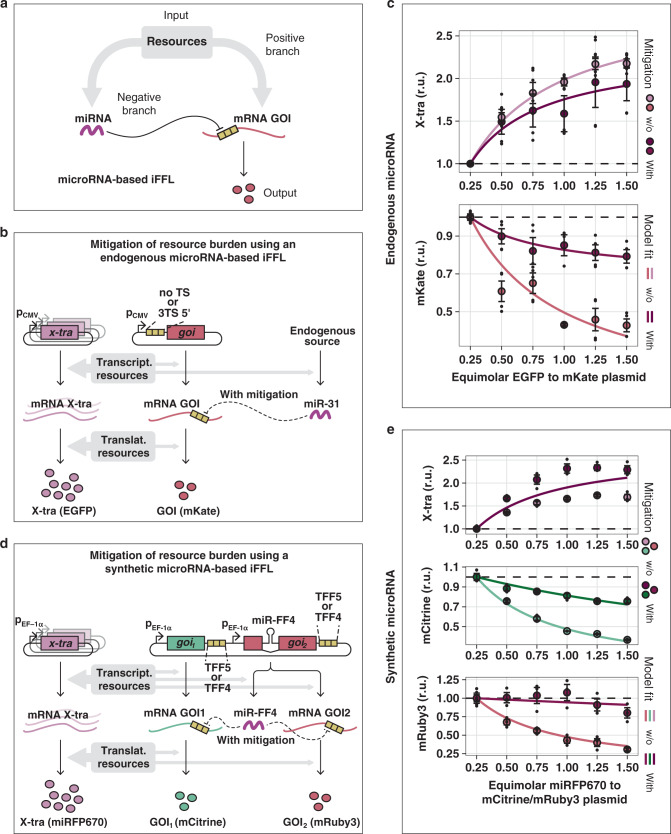
Despite recent advances in circuit engineering, the design of genetic networks in mammalian cells is still painstakingly slow and fraught with inexplicable failures. Here, we demonstrate that transiently expressed genes in mammalian cells compete for limited transcriptional and translational resources. This competition results in the coupling of otherwise independent exogenous and endogenous genes, creating a divergence between intended and actual function. Guided by a resource-aware mathematical model, we identify and engineer natural and synthetic miRNA-based incoherent feedforward loop (iFFL) circuits that mitigate gene expression burden. The implementation of these circuits features the use of endogenous miRNAs as elementary components of the engineered iFFL device, a versatile hybrid design that allows burden mitigation to be achieved across different cell-lines with minimal resource requirements. This study establishes the foundations for context-aware prediction and improvement of in vivo synthetic circuit performance, paving the way towards more rational synthetic construct design in mammalian cells.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
MIRELLA: a mathematical model explains the effect of microRNA-mediated synthetic genes regulation on intracellular resource allocation.Nucleic Acids Res. 2023 Apr 24;51(7):3452-3464. doi: 10.1093/nar/gkad151. Nucleic Acids Res. 2023. PMID: 36912077 Free PMC article.
-
Negatively Competitive Incoherent Feedforward Loops Mitigate Winner-Take-All Resource Competition.ACS Synth Biol. 2022 Dec 16;11(12):3986-3995. doi: 10.1021/acssynbio.2c00318. Epub 2022 Nov 10. ACS Synth Biol. 2022. PMID: 36355441 Free PMC article.
-
Design and Evaluation of Synthetic RNA-Based Incoherent Feed-Forward Loop Circuits.Biomolecules. 2021 Aug 10;11(8):1182. doi: 10.3390/biom11081182. Biomolecules. 2021. PMID: 34439849 Free PMC article.
-
Mammalian gene circuits with biomolecule-responsive RNA devices.Curr Opin Chem Biol. 2019 Oct;52:16-22. doi: 10.1016/j.cbpa.2019.04.013. Epub 2019 May 23. Curr Opin Chem Biol. 2019. PMID: 31129468 Review.
-
Synthetic RNA-based post-transcriptional expression control methods and genetic circuits.Adv Drug Deliv Rev. 2022 May;184:114196. doi: 10.1016/j.addr.2022.114196. Epub 2022 Mar 11. Adv Drug Deliv Rev. 2022. PMID: 35288218 Review.
Cited by
-
A genetic mammalian proportional-integral feedback control circuit for robust and precise gene regulation.Proc Natl Acad Sci U S A. 2022 Jun 14;119(24):e2122132119. doi: 10.1073/pnas.2122132119. Epub 2022 Jun 10. Proc Natl Acad Sci U S A. 2022. PMID: 35687671 Free PMC article.
-
Termination sequence between an inducible promoter and ubiquitous chromatin opening element (UCOE) reduces gene expression leakage and silencing.J Biol Eng. 2025 Apr 9;19(1):29. doi: 10.1186/s13036-025-00499-8. J Biol Eng. 2025. PMID: 40205378 Free PMC article.
-
Eukaryotic gene regulation at equilibrium, or non?Curr Opin Syst Biol. 2022 Sep;31:100435. doi: 10.1016/j.coisb.2022.100435. Epub 2022 Oct 20. Curr Opin Syst Biol. 2022. PMID: 36590072 Free PMC article.
-
Context-dependent redesign of robust synthetic gene circuits.Trends Biotechnol. 2024 Jul;42(7):895-909. doi: 10.1016/j.tibtech.2024.01.003. Epub 2024 Feb 5. Trends Biotechnol. 2024. PMID: 38320912 Free PMC article. Review.
-
Computational identification of small molecules for increased gene expression by synthetic circuits in mammalian cells.Nat Commun. 2025 Aug 4;16(1):7160. doi: 10.1038/s41467-025-62529-9. Nat Commun. 2025. PMID: 40759644 Free PMC article.
References
-
- Brinkman BM, Zuijdeest D, Kaijzel EL, Breedveld FC, Verweij CL. Relevance of the tumor necrosis factor alpha (TNF alpha) -308 promoter polymorphism in TNF alpha gene regulation. J. Inflamm. 1995;46:32–41. - PubMed
-
- Bamforth SD, et al. Cardiac malformations, adrenal agenesis, neural crest defects and exencephaly in mice lacking Cited2, a new Tfap2 co-activator. Nat. Genet. 2001;29:469–474. - PubMed
-
- Liu W, et al. Mutations in AXIN2 cause colorectal cancer with defective mismatch repair by activating β-catenin/TCF signalling. Nat. Genet. 2000;26:146–147. - PubMed
-
- Stuible M, et al. Optimization of a high-cell-density polyethylenimine transfection method for rapid protein production in CHO-EBNA1 cells. J. Biotechnol. 2018;281:39–47. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
