

Design of small molecules targeting RNA structure from sequence
- PMID: 32935689
- PMCID: PMC7707016
- DOI: 10.1039/d0cs00455c
Design of small molecules targeting RNA structure from sequence
Abstract
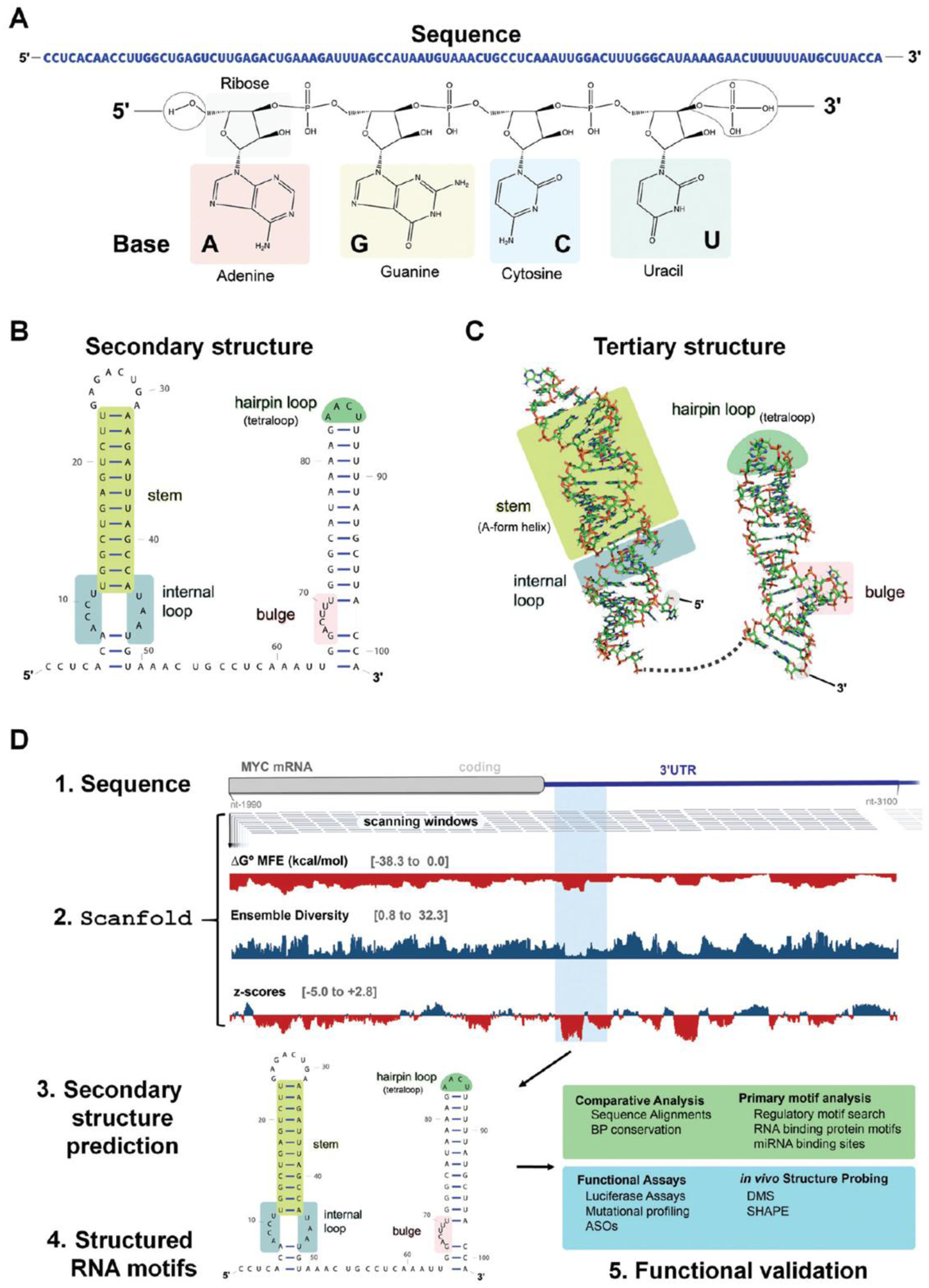
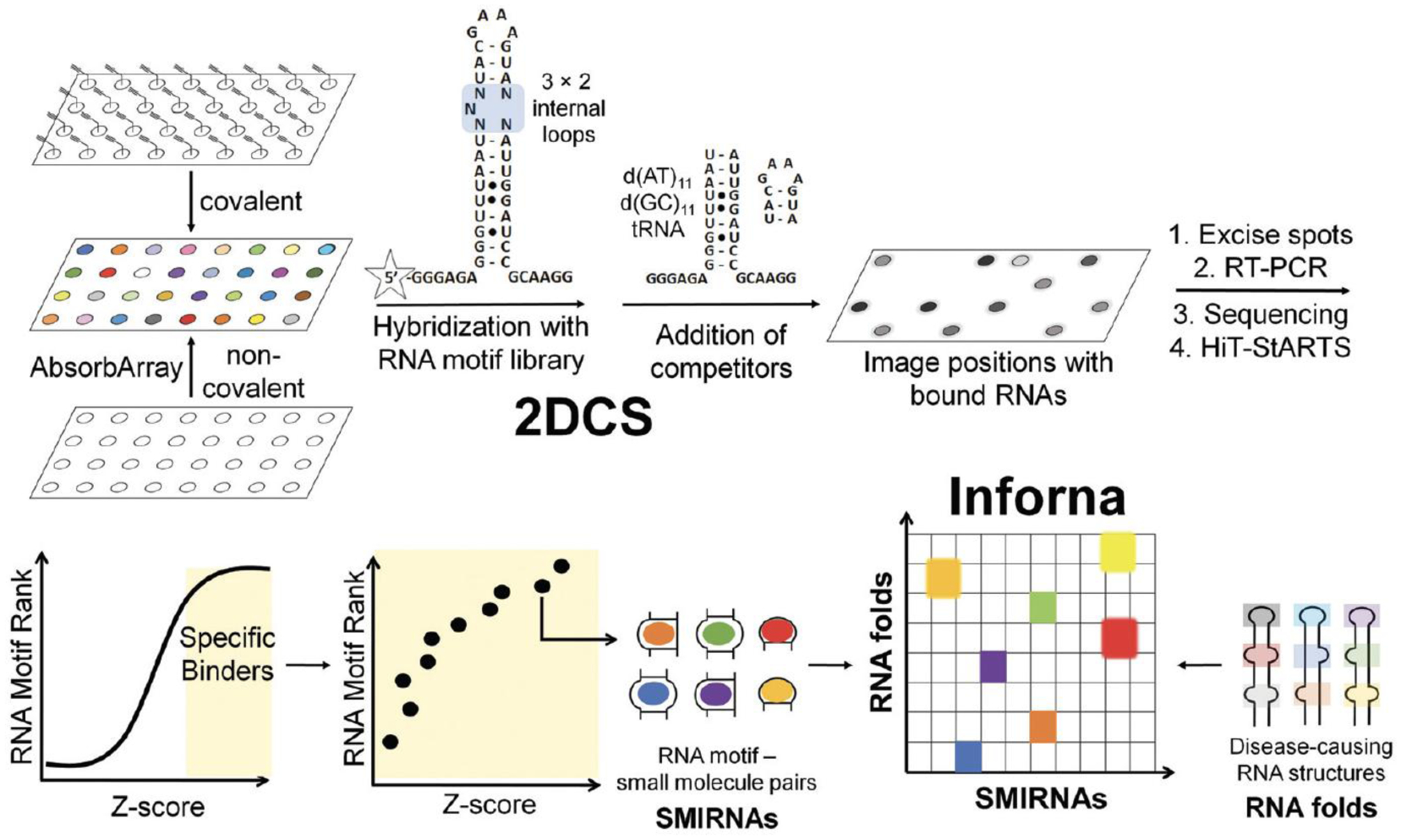
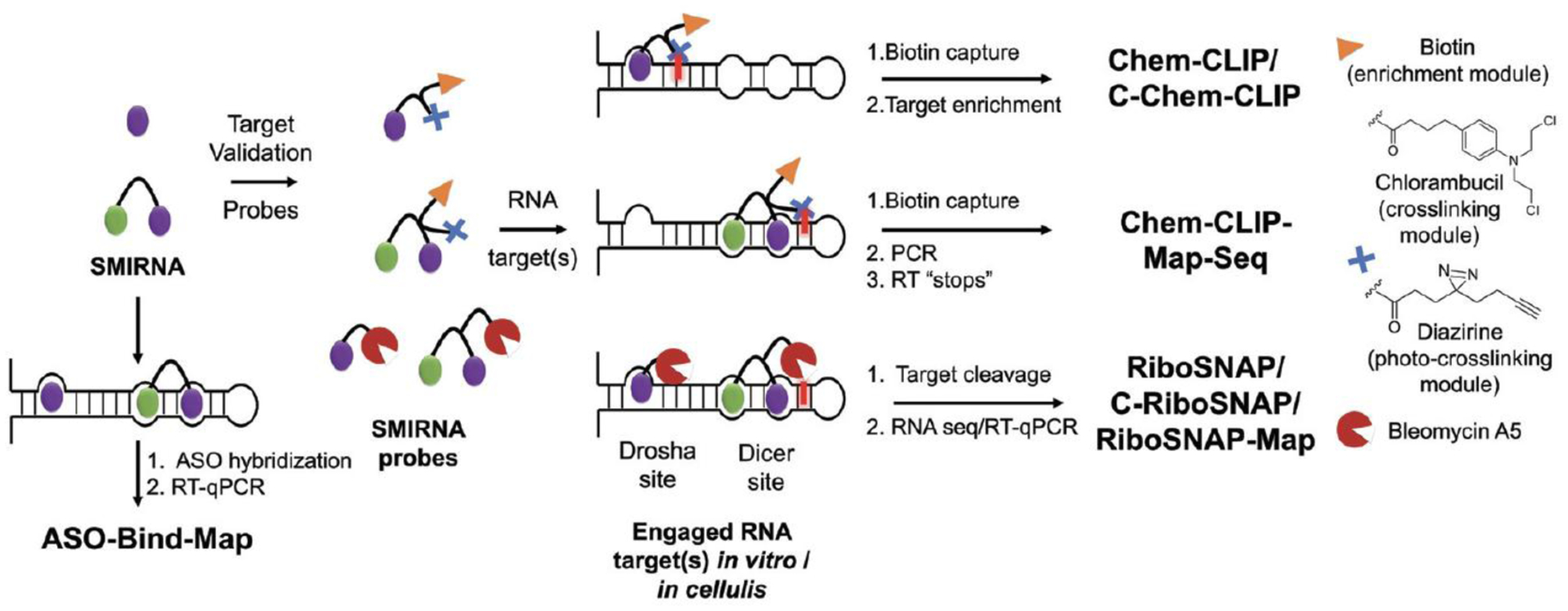
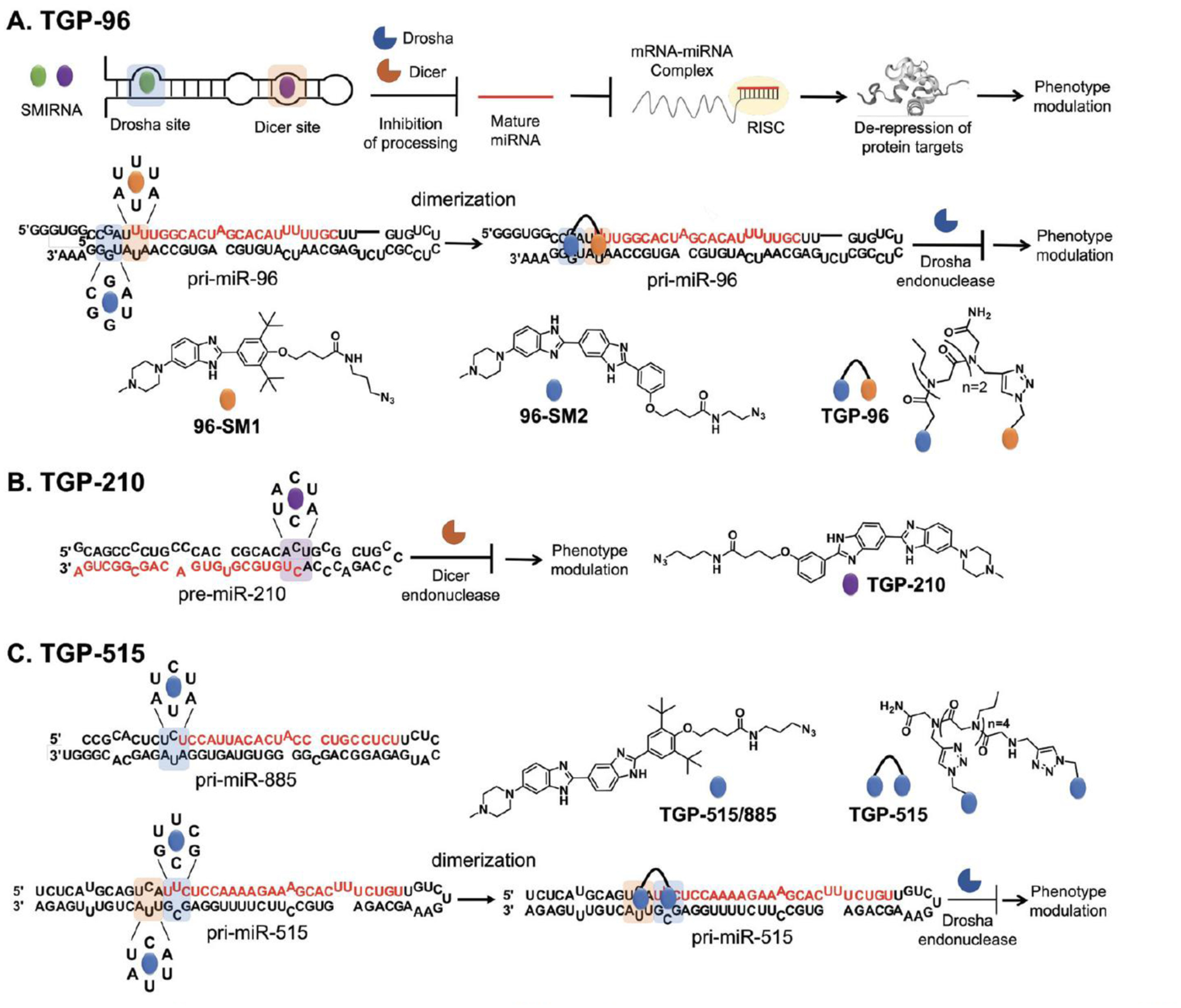
The design and discovery of small molecule medicines has largely been focused on a small number of druggable protein families. A new paradigm is emerging, however, in which small molecules exert a biological effect by interacting with RNA, both to study human disease biology and provide lead therapeutic modalities. Due to this potential for expanding target pipelines and treating a larger number of human diseases, robust platforms for the rational design and optimization of small molecules interacting with RNAs (SMIRNAs) are in high demand. This review highlights three major pillars in this area. First, the transcriptome-wide identification and validation of structured RNA elements, or motifs, within disease-causing RNAs directly from sequence is presented. Second, we provide an overview of high-throughput screening approaches to identify SMIRNAs as well as discuss the lead identification strategy, Inforna, which decodes the three-dimensional (3D) conformation of RNA motifs with small molecule binding partners, directly from sequence. An emphasis is placed on target validation methods to study the causality between modulating the RNA motif in vitro and the phenotypic outcome in cells. Third, emergent modalities that convert occupancy-driven mode of action SMIRNAs into event-driven small molecule chemical probes, such as RNA cleavers and degraders, are presented. Finally, the future of the small molecule RNA therapeutics field is discussed, as well as hurdles to overcome to develop potent and selective RNA-centric chemical probes.
Conflict of interest statement
Conflicts of interest
M. D. D. is a founder of Expansion Therapeutics.
Figures
Similar articles
-
Identifying and validating small molecules interacting with RNA (SMIRNAs).Methods Enzymol. 2019;623:45-66. doi: 10.1016/bs.mie.2019.04.027. Epub 2019 May 15. Methods Enzymol. 2019. PMID: 31239057 Free PMC article.
-
Targeting RNA with Small Molecules To Capture Opportunities at the Intersection of Chemistry, Biology, and Medicine.J Am Chem Soc. 2019 May 1;141(17):6776-6790. doi: 10.1021/jacs.8b13419. Epub 2019 Apr 19. J Am Chem Soc. 2019. PMID: 30896935 Free PMC article.
-
Rational Design of Small Molecules Targeting Oncogenic Noncoding RNAs from Sequence.Acc Chem Res. 2016 Dec 20;49(12):2698-2704. doi: 10.1021/acs.accounts.6b00326. Epub 2016 Nov 22. Acc Chem Res. 2016. PMID: 27993012 Free PMC article.
-
RNA as a small molecule druggable target.Bioorg Med Chem Lett. 2017 Dec 1;27(23):5083-5088. doi: 10.1016/j.bmcl.2017.10.052. Epub 2017 Oct 23. Bioorg Med Chem Lett. 2017. PMID: 29097169 Review.
-
Unveiling the druggable RNA targets and small molecule therapeutics.Bioorg Med Chem. 2019 May 15;27(10):2149-2165. doi: 10.1016/j.bmc.2019.03.057. Epub 2019 Mar 30. Bioorg Med Chem. 2019. PMID: 30981606 Free PMC article. Review.
Cited by
-
Small molecule recognition of disease-relevant RNA structures.Chem Soc Rev. 2020 Oct 5;49(19):7167-7199. doi: 10.1039/d0cs00560f. Chem Soc Rev. 2020. PMID: 32975549 Free PMC article. Review.
-
A map of the SARS-CoV-2 RNA structurome.NAR Genom Bioinform. 2021 May 22;3(2):lqab043. doi: 10.1093/nargab/lqab043. eCollection 2021 Jun. NAR Genom Bioinform. 2021. PMID: 34046592 Free PMC article.
-
Rational guide RNA engineering for small-molecule control of CRISPR/Cas9 and gene editing.Nucleic Acids Res. 2022 May 6;50(8):4769-4783. doi: 10.1093/nar/gkac255. Nucleic Acids Res. 2022. PMID: 35446403 Free PMC article.
-
MicroRNAs in the Regulation of Solute Carrier Proteins Behind Xenobiotic and Nutrient Transport in Cells.Front Mol Biosci. 2022 Jun 9;9:893846. doi: 10.3389/fmolb.2022.893846. eCollection 2022. Front Mol Biosci. 2022. PMID: 35755805 Free PMC article. Review.
-
RNA-Targeting Splicing Modifiers: Drug Development and Screening Assays.Molecules. 2021 Apr 14;26(8):2263. doi: 10.3390/molecules26082263. Molecules. 2021. PMID: 33919699 Free PMC article. Review.
References
-
- Hendrix DK, Brenner SE and Holbrook SR, Q. Rev. Biophys, 2005, 38, 221–243. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources