

Innate Lymphoid Cell Activation and Sustained Depletion in Blood and Tissue of Children Infected with HIV from Birth Despite Antiretroviral Therapy
- PMID: 32937142
- PMCID: PMC7495043
- DOI: 10.1016/j.celrep.2020.108153
Innate Lymphoid Cell Activation and Sustained Depletion in Blood and Tissue of Children Infected with HIV from Birth Despite Antiretroviral Therapy
Abstract
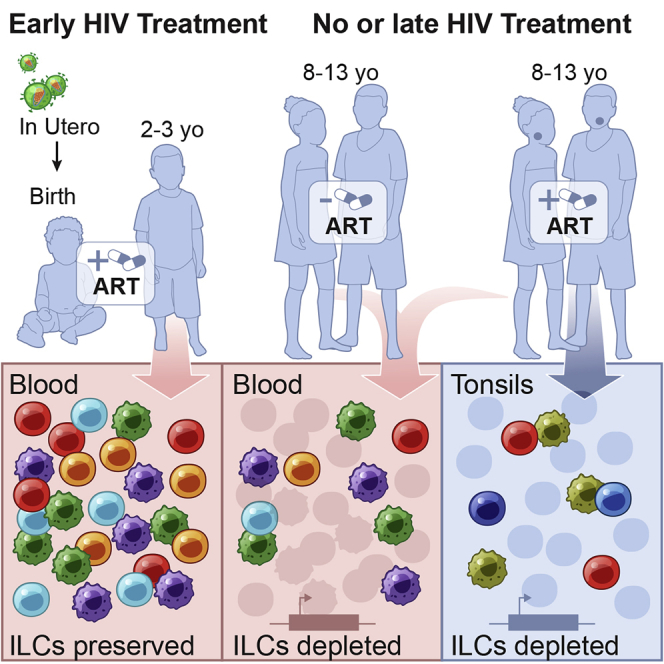
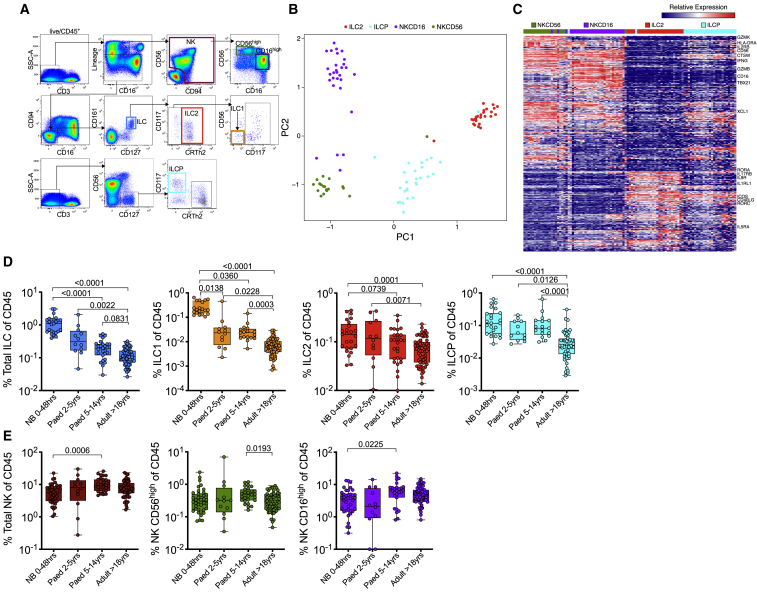
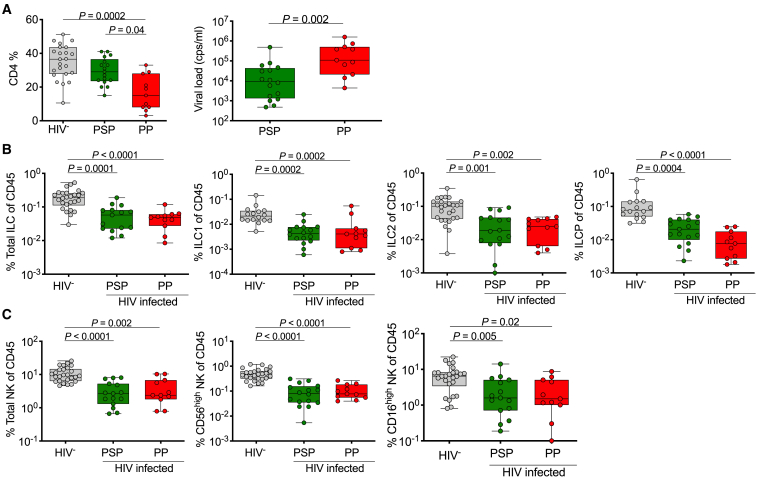
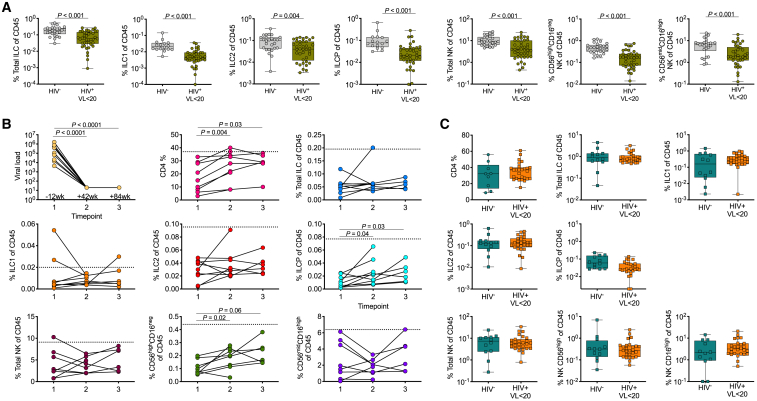
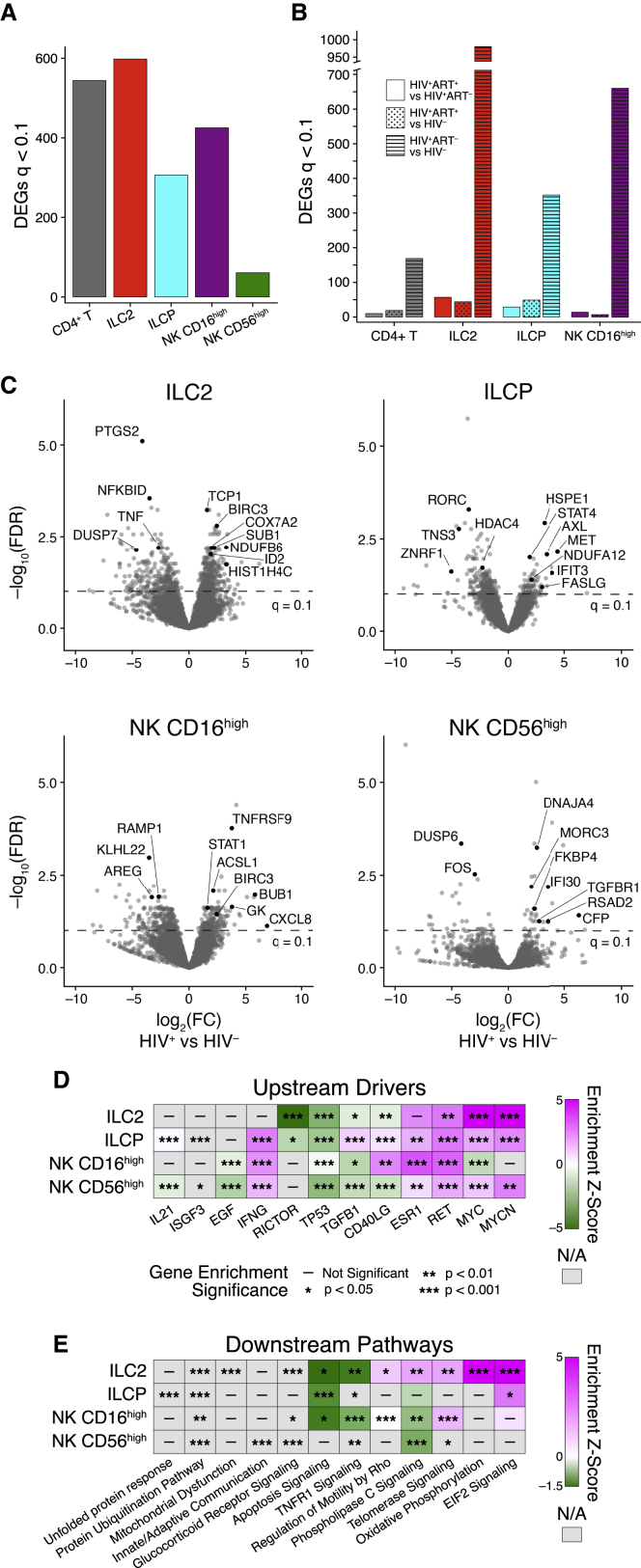
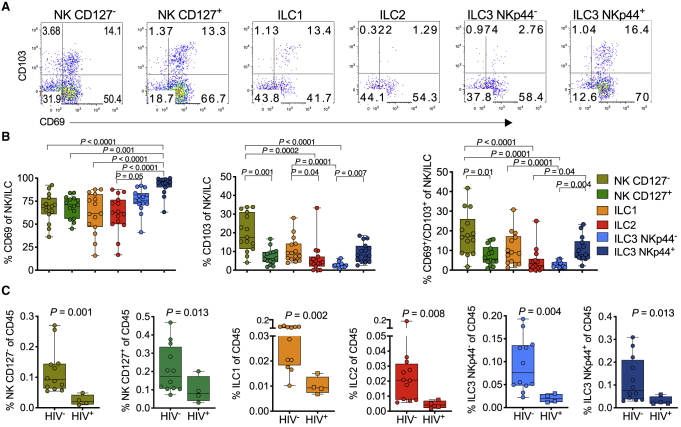
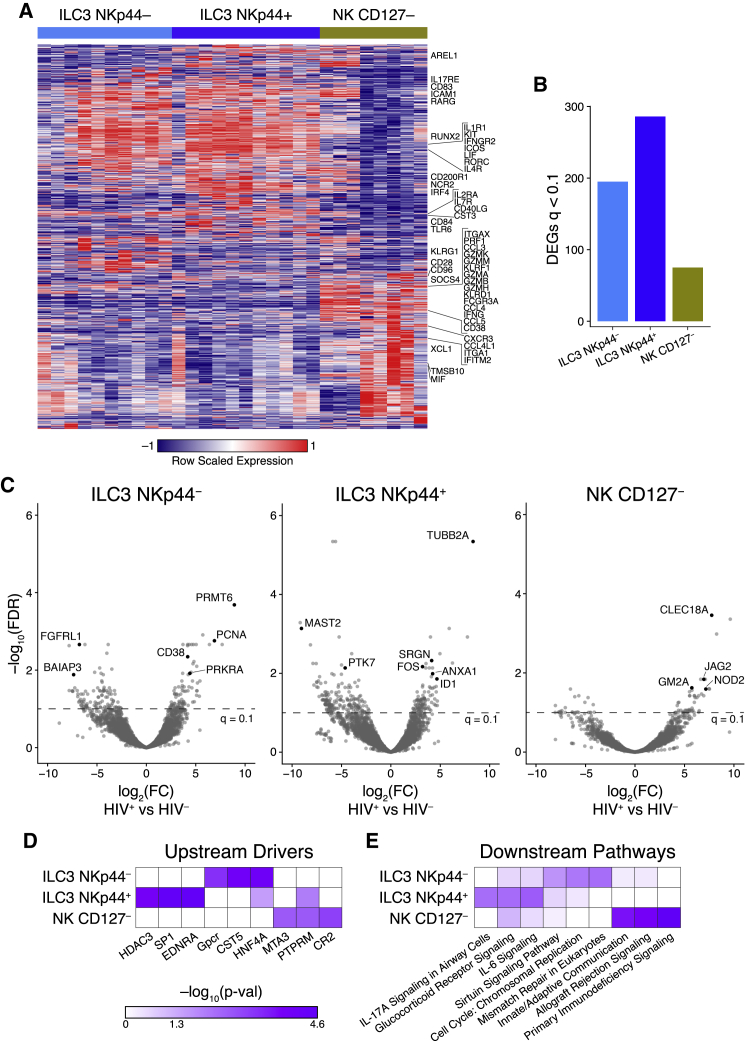
Innate lymphoid cells (ILCs) are important for response to infection and for immune development in early life. HIV infection in adults depletes circulating ILCs, but the impact on children infected from birth remains unknown. We study vertically HIV-infected children from birth to adulthood and find severe and persistent depletion of all circulating ILCs that, unlike CD4+ T cells, are not restored by long-term antiretroviral therapy unless initiated at birth. Remaining ILCs upregulate genes associated with cellular activation and metabolic perturbation. Unlike HIV-infected adults, ILCs are also profoundly depleted in tonsils of vertically infected children. Transcriptional profiling of remaining ILCs reveals ongoing cell-type-specific activity despite antiretroviral therapy. Collectively, these data suggest an important and ongoing role for ILCs in lymphoid tissue of HIV-infected children from birth, where persistent depletion and sustained transcriptional activity are likely to have long-term immune consequences that merit further investigation.
Keywords: HIV; ILCs; NK cells; pediatric infection; tonsil; vertical transmission.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Arcone R., Arpaia G., Ruoppolo M., Malorni A., Pucci P., Marino G., Ialenti A., Di Rosa M., Ciliberto G. Structural characterization of a biologically active human lipocortin 1 expressed in Escherichia coli. Eur. J. Biochem. 1993;211:347–355. - PubMed
-
- Azzoni L., Rutstein R.M., Chehimi J., Farabaugh M.A., Nowmos A., Montaner L.J. Dendritic and natural killer cell subsets associated with stable or declining CD4+ cell counts in treated HIV-1-infected children. J. Infect. Dis. 2005;191:1451–1459. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
