

Dietary Regulation of Immunity
- PMID: 32937152
- PMCID: PMC7491384
- DOI: 10.1016/j.immuni.2020.08.013
Dietary Regulation of Immunity
Abstract
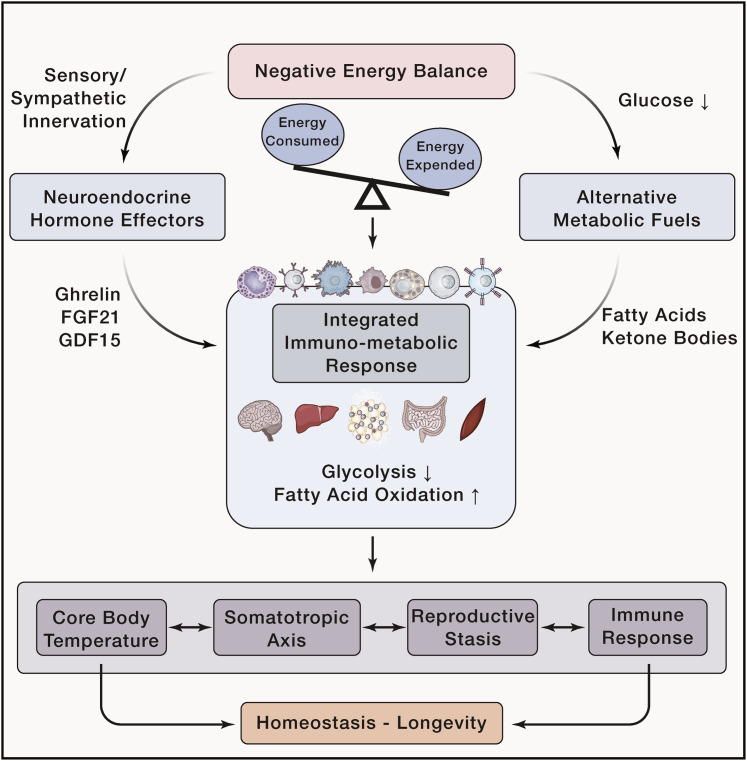
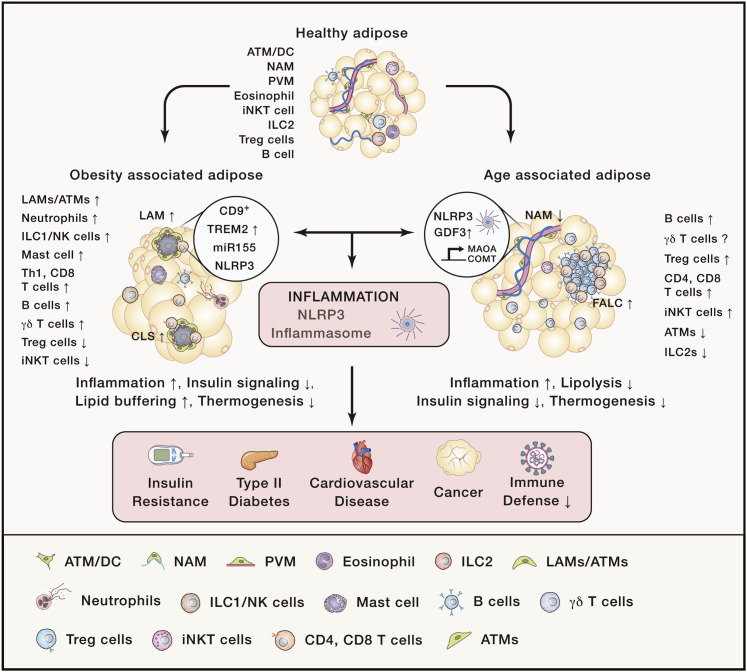
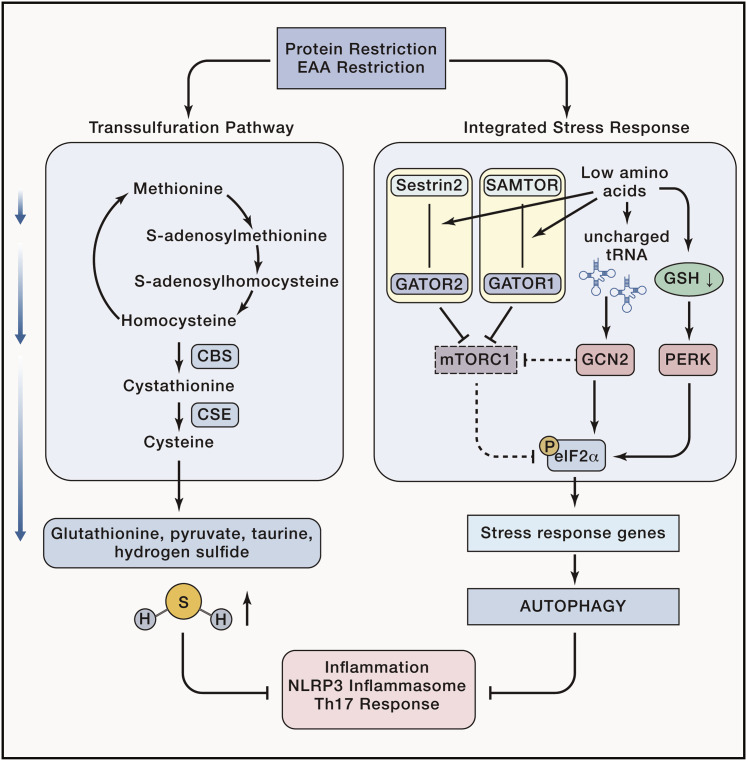
Integrated immunometabolic responses link dietary intake, energy utilization, and storage to immune regulation of tissue function and is therefore essential for the maintenance and restoration of homeostasis. Adipose-resident leukocytes have non-traditional immunological functions that regulate organismal metabolism by controlling insulin action, lipolysis, and mitochondrial respiration to control the usage of substrates for production of heat versus ATP. Energetically expensive vital functions such as immunological responses might have thus evolved to respond accordingly to dietary surplus and deficit of macronutrient intake. Here, we review the interaction of dietary intake of macronutrients and their metabolism with the immune system. We discuss immunometabolic checkpoints that promote healthspan and highlight how dietary fate and regulation of glucose, fat, and protein metabolism might affect immunity.
Copyright © 2020 Elsevier Inc. All rights reserved.
Figures
References
-
- Arkan M.C., Hevener A.L., Greten F.R., Maeda S., Li Z.W., Long J.M., Wynshaw-Boris A., Poli G., Olefsky J., Karin M. IKK-beta links inflammation to obesity-induced insulin resistance. Nat. Med. 2005;11:191–198. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
