

Integrating Mathematical Modeling with High-Throughput Imaging Explains How Polyploid Populations Behave in Nutrient-Sparse Environments
- PMID: 32938640
- PMCID: PMC7671819
- DOI: 10.1158/0008-5472.CAN-20-1231
Integrating Mathematical Modeling with High-Throughput Imaging Explains How Polyploid Populations Behave in Nutrient-Sparse Environments
Abstract
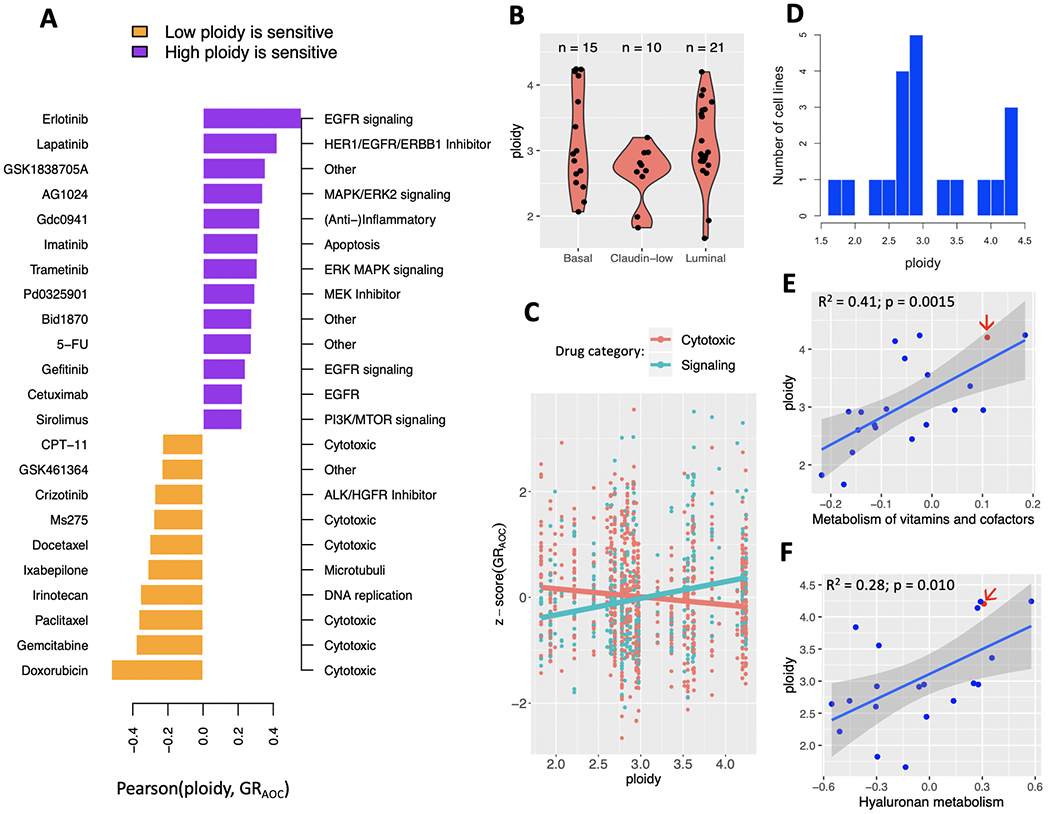
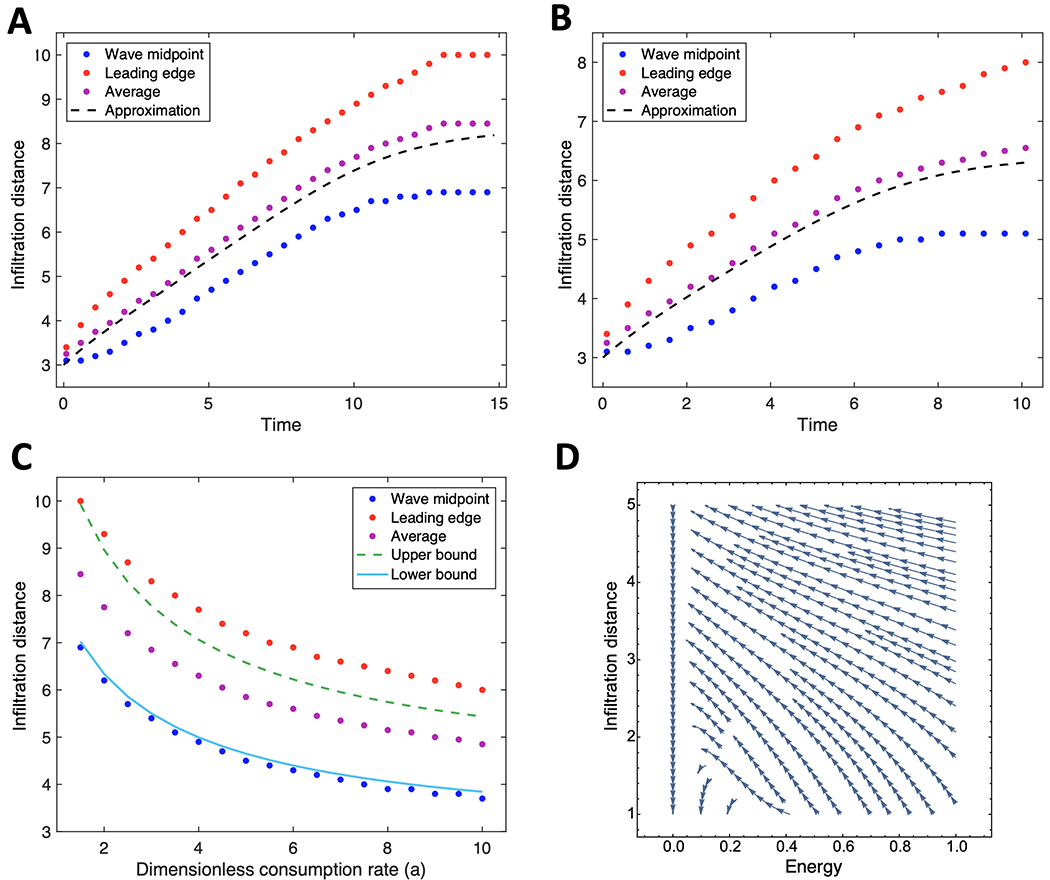
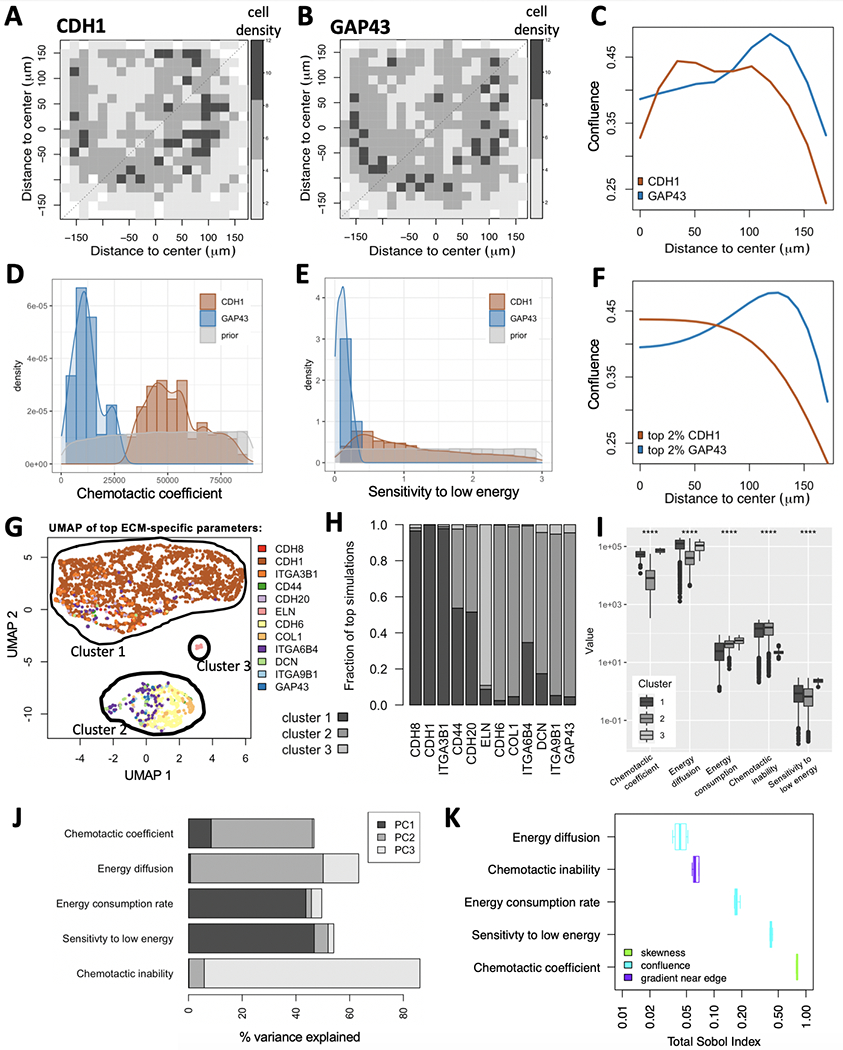
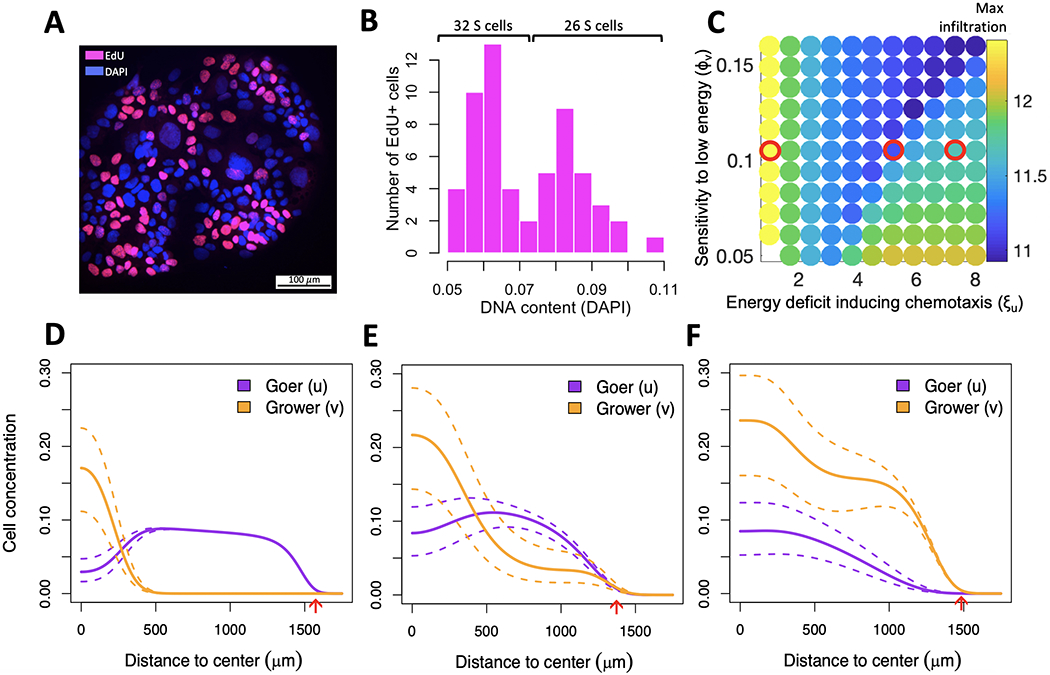
Breast cancer progresses in a multistep process from primary tumor growth and stroma invasion to metastasis. Nutrient-limiting environments promote chemotaxis with aggressive morphologies characteristic of invasion. It is unknown how coexisting cells differ in their response to nutrient limitations and how this impacts invasion of the metapopulation as a whole. In this study, we integrate mathematical modeling with microenvironmental perturbation data to investigate invasion in nutrient-limiting environments inhabited by one or two cancer cell subpopulations. Subpopulations were defined by their energy efficiency and chemotactic ability. Invasion distance traveled by a homogeneous population was estimated. For heterogeneous populations, results suggest that an imbalance between nutrient efficacy and chemotactic superiority accelerates invasion. Such imbalance will spatially segregate the two populations and only one type will dominate at the invasion front. Only if these two phenotypes are balanced, the two subpopulations compete for the same space, which decelerates invasion. We investigate ploidy as a candidate biomarker of this phenotypic heterogeneity and discuss its potential to inform the dose of mTOR inhibitors (mTOR-I) that can inhibit chemotaxis just enough to facilitate such competition. SIGNIFICANCE: This study identifies the double-edged sword of high ploidy as a prerequisite to personalize combination therapies with cytotoxic drugs and inhibitors of signal transduction pathways such as mTOR-Is. GRAPHICAL ABSTRACT: http://cancerres.aacrjournals.org/content/canres/80/22/5109/F1.large.jpg.
©2020 American Association for Cancer Research.
Conflict of interest statement
Conflict of Interest
The authors declare no potential conflicts of interest.
Figures
References
-
- Angelova M, Mlecnik B, Vasaturo A, Bindea G, Fredriksen T, Lafontaine L, et al. Evolution of Metastases in Space and Time under Immune Selection. Cell. 2018;175:751–765.e16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
