

The Enzymatic Activity of the nsp14 Exoribonuclease Is Critical for Replication of MERS-CoV and SARS-CoV-2
- PMID: 32938769
- PMCID: PMC7654266
- DOI: 10.1128/JVI.01246-20
The Enzymatic Activity of the nsp14 Exoribonuclease Is Critical for Replication of MERS-CoV and SARS-CoV-2
Abstract
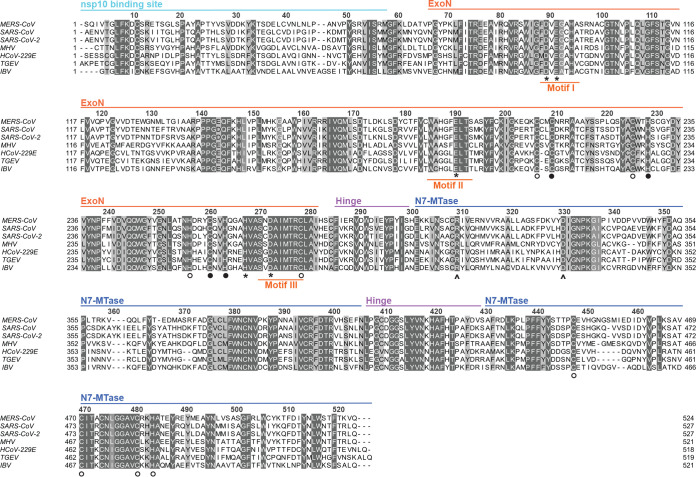
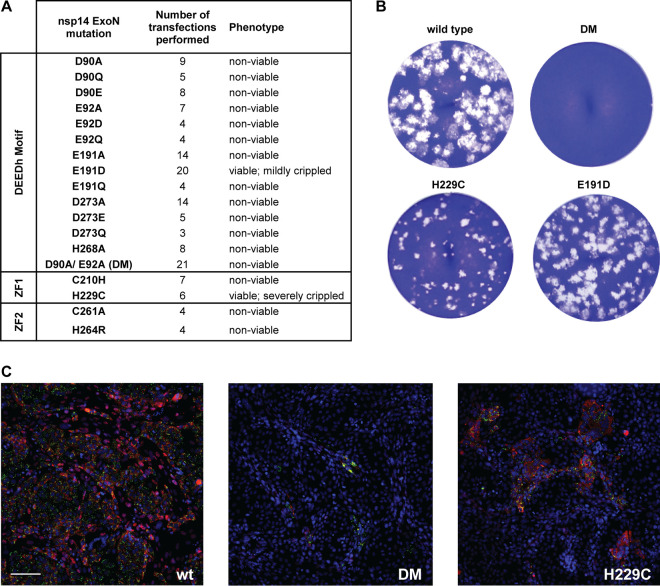
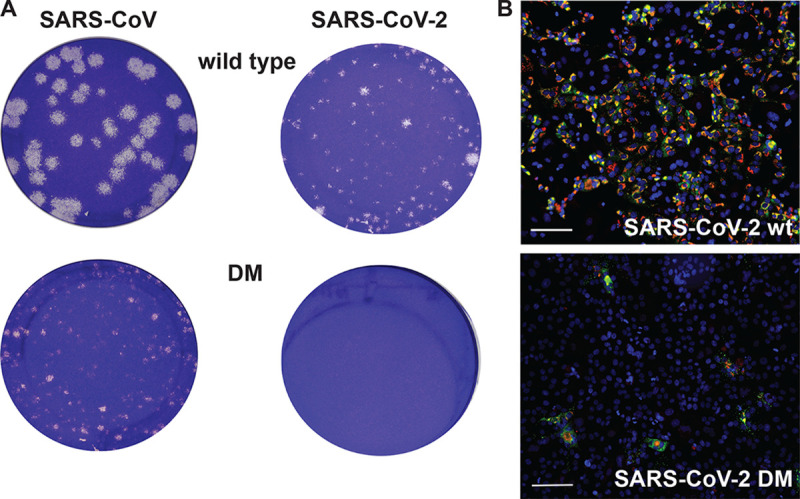
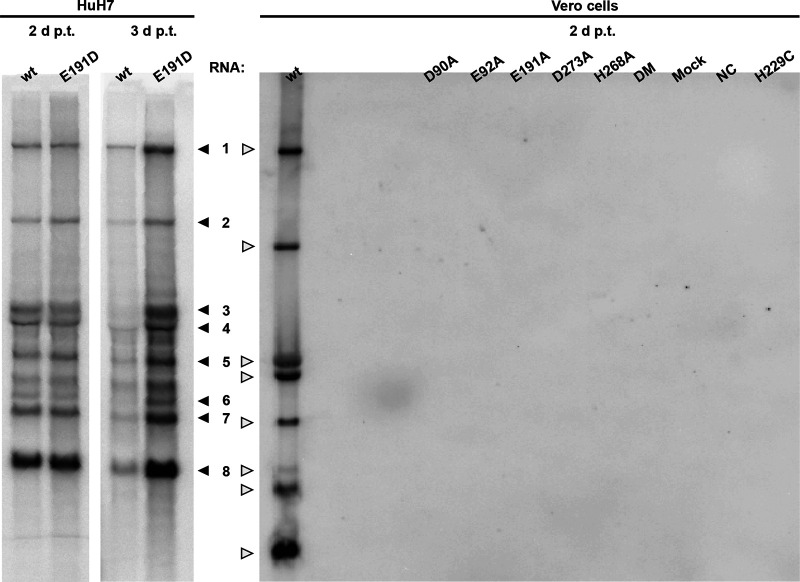
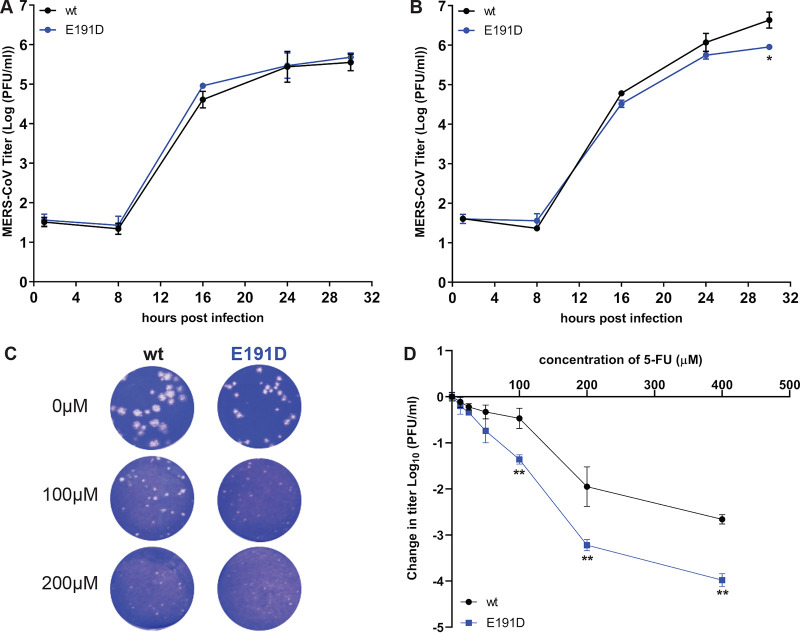
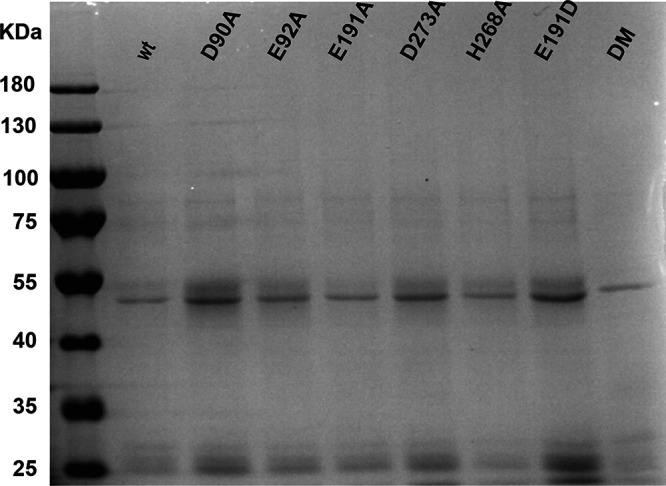
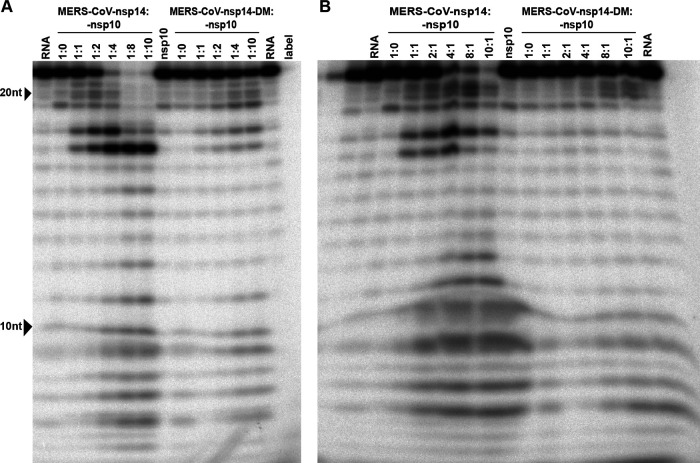
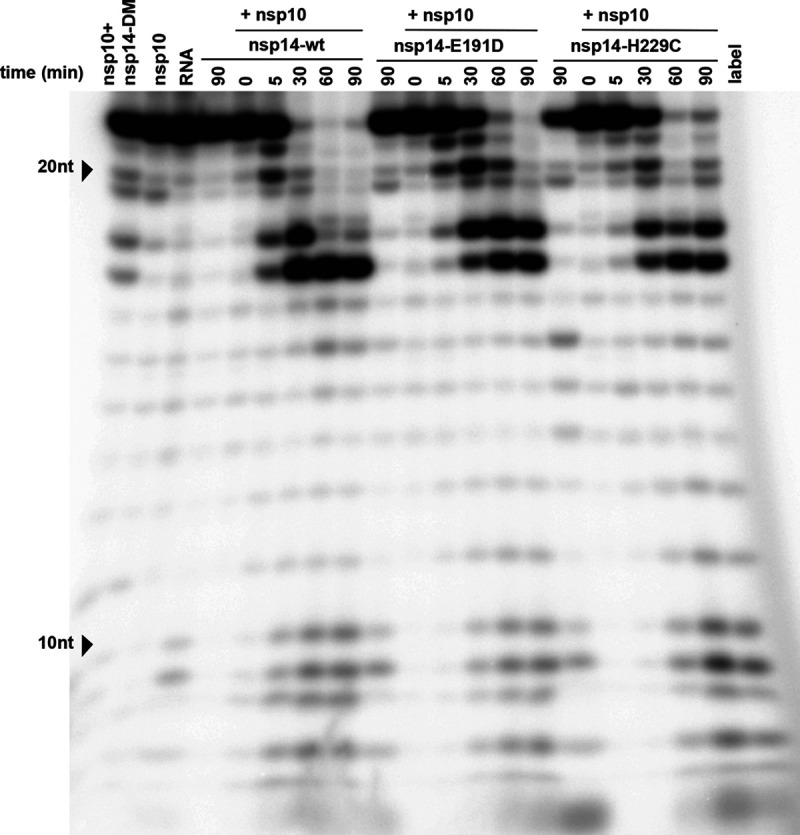
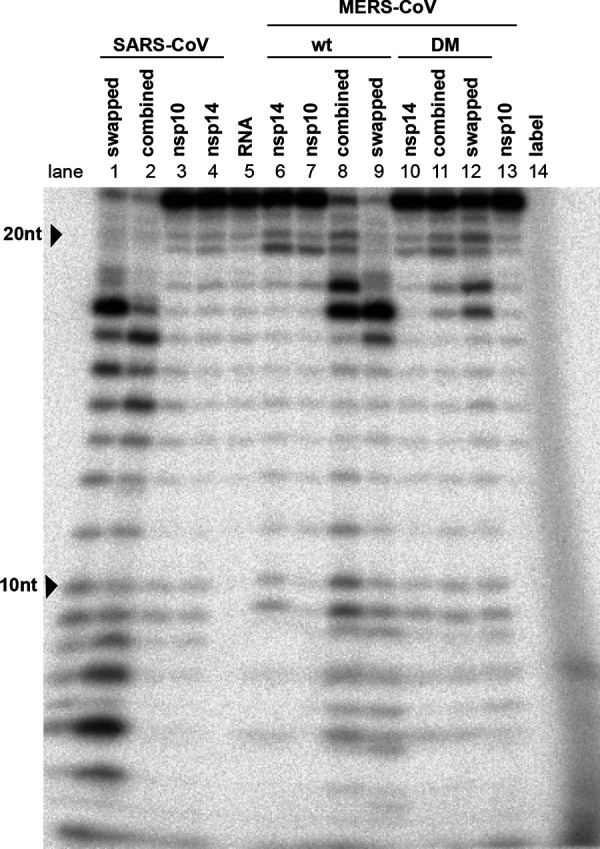
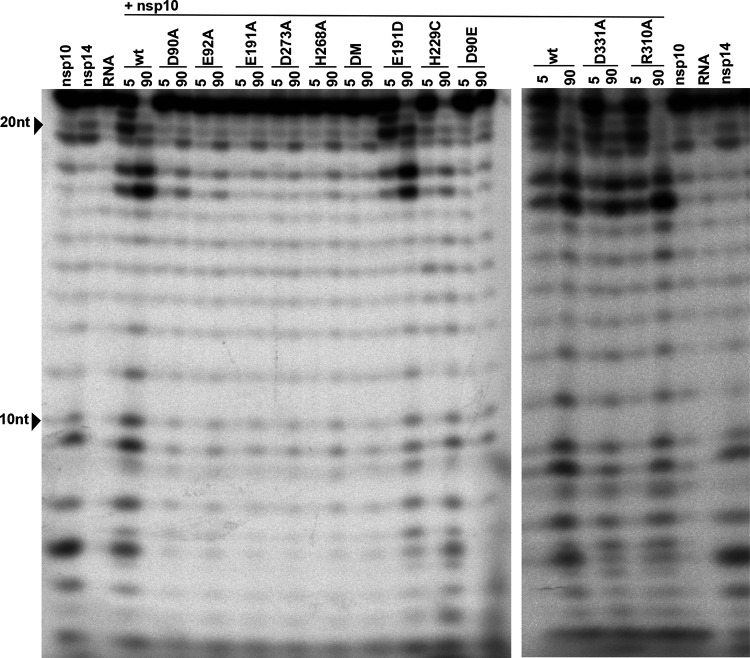
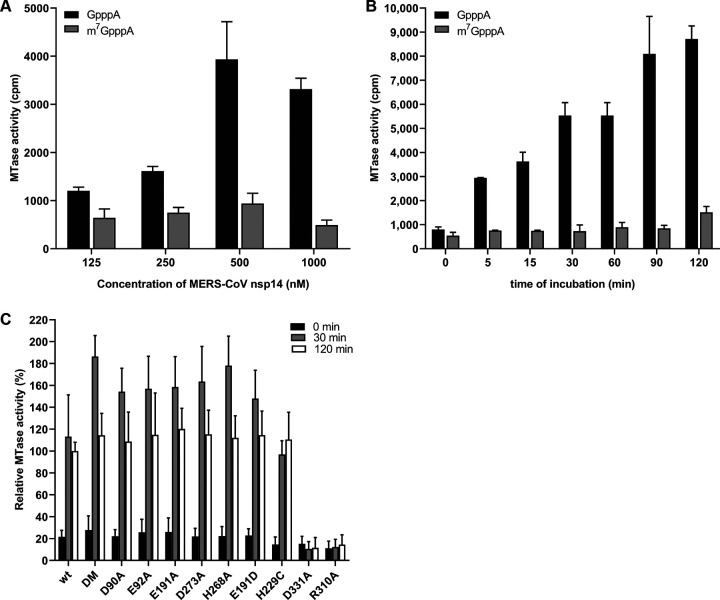
Coronaviruses (CoVs) stand out for their large RNA genome and complex RNA-synthesizing machinery comprising 16 nonstructural proteins (nsps). The bifunctional nsp14 contains 3'-to-5' exoribonuclease (ExoN) and guanine-N7-methyltransferase (N7-MTase) domains. While the latter presumably supports mRNA capping, ExoN is thought to mediate proofreading during genome replication. In line with such a role, ExoN knockout mutants of mouse hepatitis virus (MHV) and severe acute respiratory syndrome coronavirus (SARS-CoV) were previously reported to have crippled but viable hypermutation phenotypes. Remarkably, using reverse genetics, a large set of corresponding ExoN knockout mutations has now been found to be lethal for another betacoronavirus, Middle East respiratory syndrome coronavirus (MERS-CoV). For 13 mutants, viral progeny could not be recovered, unless-as happened occasionally-reversion had first occurred. Only a single mutant was viable, likely because its E191D substitution is highly conservative. Remarkably, a SARS-CoV-2 ExoN knockout mutant was found to be unable to replicate, resembling observations previously made for alpha- and gammacoronaviruses, but starkly contrasting with the documented phenotype of ExoN knockout mutants of the closely related SARS-CoV. Subsequently, we established in vitro assays with purified recombinant MERS-CoV nsp14 to monitor its ExoN and N7-MTase activities. All ExoN knockout mutations that proved lethal in reverse genetics were found to severely decrease ExoN activity while not affecting N7-MTase activity. Our study strongly suggests that CoV nsp14 ExoN has an additional function, which apparently is critical for primary viral RNA synthesis and thus differs from the proofreading function that, based on previous MHV and SARS-CoV studies, was proposed to boost longer-term replication fidelity.IMPORTANCE The bifunctional nsp14 subunit of the coronavirus replicase contains 3'-to-5' exoribonuclease (ExoN) and guanine-N7-methyltransferase domains. For the betacoronaviruses MHV and SARS-CoV, ExoN was reported to promote the fidelity of genome replication, presumably by mediating a form of proofreading. For these viruses, ExoN knockout mutants are viable while displaying an increased mutation frequency. Strikingly, we have now established that the equivalent ExoN knockout mutants of two other betacoronaviruses, MERS-CoV and SARS-CoV-2, are nonviable, suggesting an additional and critical ExoN function in their replication. This is remarkable in light of the very limited genetic distance between SARS-CoV and SARS-CoV-2, which is highlighted, for example, by 95% amino acid sequence identity in their nsp14 sequences. For (recombinant) MERS-CoV nsp14, both its enzymatic activities were evaluated using newly developed in vitro assays that can be used to characterize these key replicative enzymes in more detail and explore their potential as target for antiviral drug development.
Keywords: RNA synthesis; guanine-N7-methyltransferase; nidovirus; nonstructural protein; proofreading; replicase; reverse genetics.
Copyright © 2020 Ogando et al.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
