

Heparan sulfate proteoglycan-mediated dynamin-dependent transport of neural stem cell exosomes in an in vitro blood-brain barrier model
- PMID: 32939863
- PMCID: PMC7891616
- DOI: 10.1111/ejn.14974
Heparan sulfate proteoglycan-mediated dynamin-dependent transport of neural stem cell exosomes in an in vitro blood-brain barrier model
Abstract
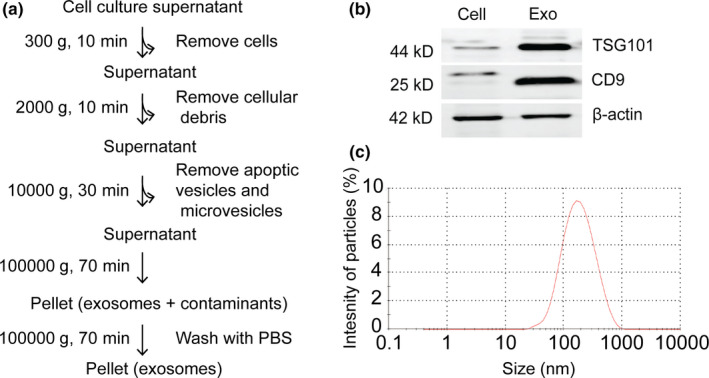
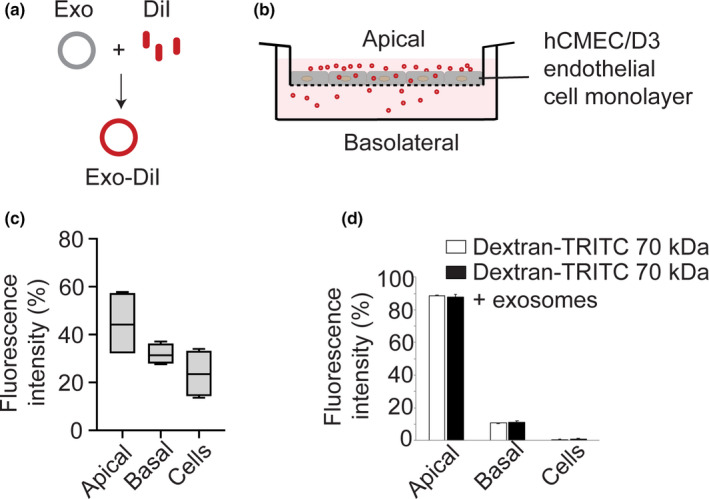
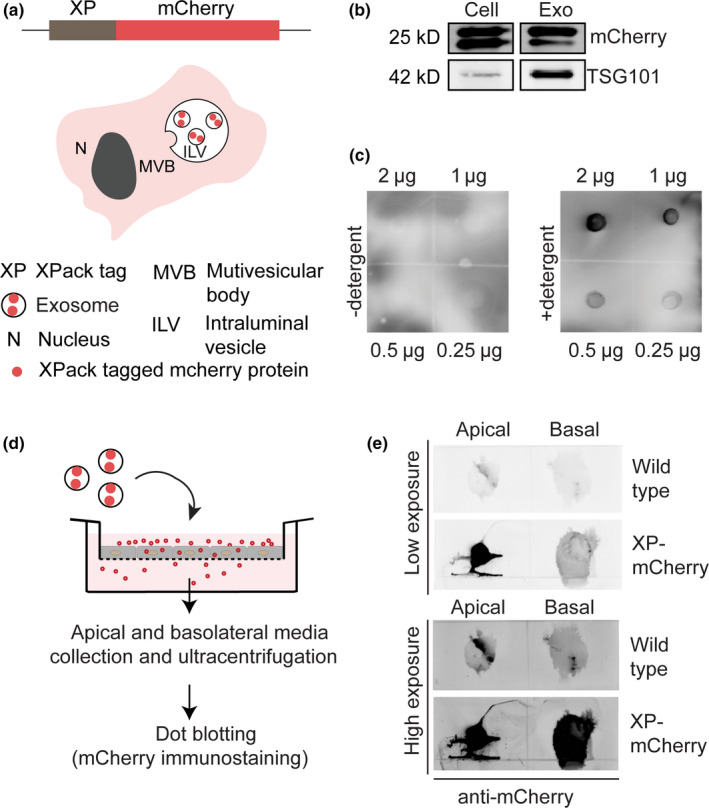
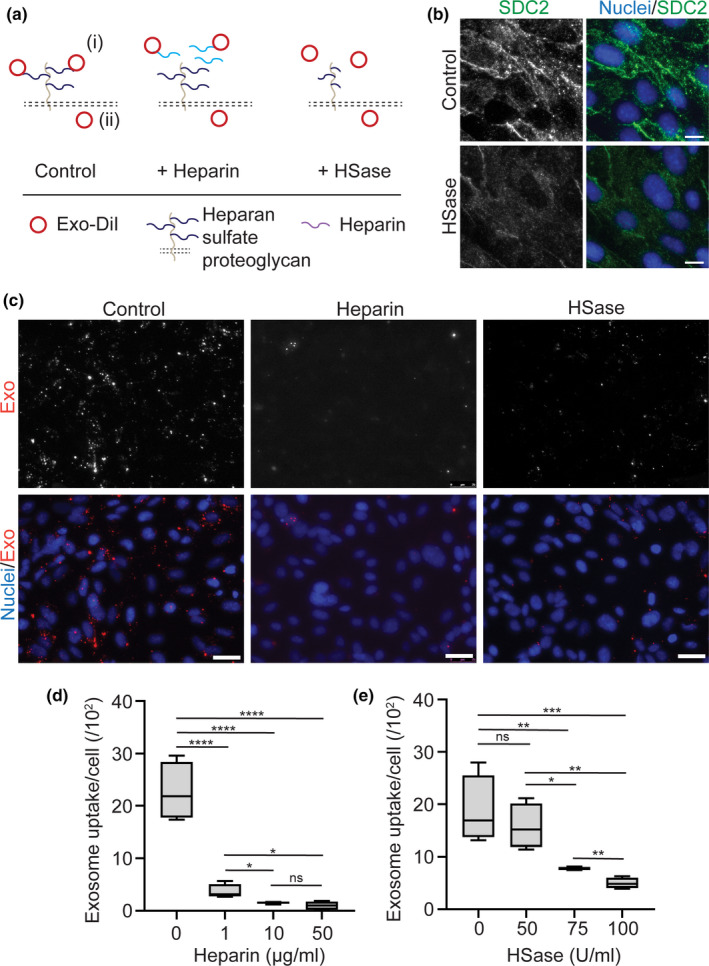
Drug delivery to the brain is greatly hampered by the presence of the blood-brain barrier (BBB) which tightly regulates the passage of molecules from blood to brain and vice versa. Nanocarriers, in which drugs can be encapsulated, can move across the blood-brain barrier (BBB) via the process of transcytosis, thus showing promise to improve drug delivery to the brain. Here, we demonstrate the use of natural nanovesicles, that is, exosomes, derived from C17.2 neural stem cells (NSCs) to efficiently carry a protein cargo across an in vitro BBB model consisting of human brain microvascular endothelial cells. We show that the exosomes are primarily taken up in brain endothelial cells via endocytosis, while heparan sulfate proteoglycans (HSPGs) act as receptors. Taken together, our data support the view that NSC exosomes may act as biological nanocarriers for efficient passage across the BBB. Nanomedicines that target HSPGs may improve their binding to brain endothelial cells and, possibly, show subsequent transcytosis across the BBB.
Keywords: blood-brain barrier; cargo; endothelial cell; exosomes; extracellular vesicles; heparan sulfate proteoglycans; nanocarriers; transcytosis.
© 2020 The Authors. European Journal of Neuroscience published by Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Figures

Similar articles
-
Transport of Extracellular Vesicles across the Blood-Brain Barrier: Brain Pharmacokinetics and Effects of Inflammation.Int J Mol Sci. 2020 Jun 21;21(12):4407. doi: 10.3390/ijms21124407. Int J Mol Sci. 2020. PMID: 32575812 Free PMC article.
-
Tumor-Derived Extracellular Vesicles Breach the Intact Blood-Brain Barrier via Transcytosis.ACS Nano. 2019 Dec 24;13(12):13853-13865. doi: 10.1021/acsnano.9b04397. Epub 2019 Sep 10. ACS Nano. 2019. PMID: 31479239 Free PMC article.
-
A filter-free blood-brain barrier model to quantitatively study transendothelial delivery of nanoparticles by fluorescence spectroscopy.J Control Release. 2018 Nov 10;289:14-22. doi: 10.1016/j.jconrel.2018.09.015. Epub 2018 Sep 20. J Control Release. 2018. PMID: 30243824
-
A Brief History of Adherons: The Discovery of Brain Exosomes.Int J Mol Sci. 2020 Oct 16;21(20):7673. doi: 10.3390/ijms21207673. Int J Mol Sci. 2020. PMID: 33081326 Free PMC article. Review.
-
Heparan sulfate proteoglycan as a cell-surface endocytosis receptor.Matrix Biol. 2014 Apr;35:51-5. doi: 10.1016/j.matbio.2013.10.004. Epub 2013 Oct 18. Matrix Biol. 2014. PMID: 24145152 Review.
Cited by
-
The Role of BAR Proteins and the Glycocalyx in Brain Endothelium Transcytosis.Cells. 2020 Dec 14;9(12):2685. doi: 10.3390/cells9122685. Cells. 2020. PMID: 33327645 Free PMC article. Review.
-
Exosomes: the next-generation therapeutic platform for ischemic stroke.Neural Regen Res. 2025 May 1;20(5):1221-1235. doi: 10.4103/NRR.NRR-D-23-02051. Epub 2024 May 13. Neural Regen Res. 2025. PMID: 39075892 Free PMC article.
-
Genome-wide forward genetic screening to identify receptors and proteins mediating nanoparticle uptake and intracellular processing.Nat Nanotechnol. 2024 Jul;19(7):1022-1031. doi: 10.1038/s41565-024-01629-x. Epub 2024 Mar 19. Nat Nanotechnol. 2024. PMID: 38504023
-
Glioma-Derived Exosomes and Their Application as Drug Nanoparticles.Int J Mol Sci. 2024 Nov 21;25(23):12524. doi: 10.3390/ijms252312524. Int J Mol Sci. 2024. PMID: 39684236 Free PMC article. Review.
-
Unraveling the Multifaceted Roles of Extracellular Vesicles: Insights into Biology, Pharmacology, and Pharmaceutical Applications for Drug Delivery.Int J Mol Sci. 2023 Dec 29;25(1):485. doi: 10.3390/ijms25010485. Int J Mol Sci. 2023. PMID: 38203656 Free PMC article. Review.
References
-
- Abbott, N. J. , & Romero, I. A. (1996). Transporting therapeutics across the blood‐brain barrier. Molecular Medicine Today, 2, 106–113. - PubMed
-
- Aboody, K. S. , Najbauer, J. , & Danks, M. K. (2008). Stem and progenitor cell‐mediated tumor selective gene therapy. Gene Therapy, 15, 739–752. - PubMed
-
- Alpaugh, M. , Galleguillos, D. , Forero, J. , Morales, L. C. , Lackey, S. W. , Kar, P. , Di Pardo, A. , Holt, A. , Kerr, B. J. , Todd, K. G. , Baker, G. B. , Fouad, K. , & Sipione, S. (2017). Disease‐modifying effects of ganglioside GM1 in Huntington's disease models. EMBO Molecular Medicine, 9, 1537–1557. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
