

Topological constraints in early multicellularity favor reproductive division of labor
- PMID: 32940598
- PMCID: PMC7609046
- DOI: 10.7554/eLife.54348
Topological constraints in early multicellularity favor reproductive division of labor
Abstract
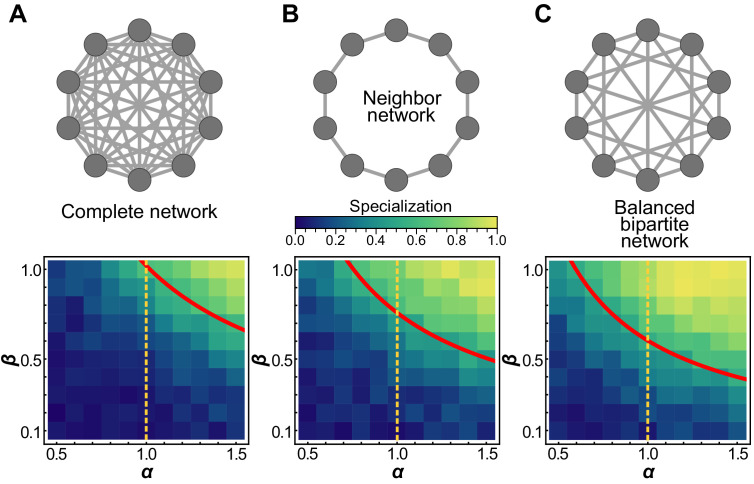
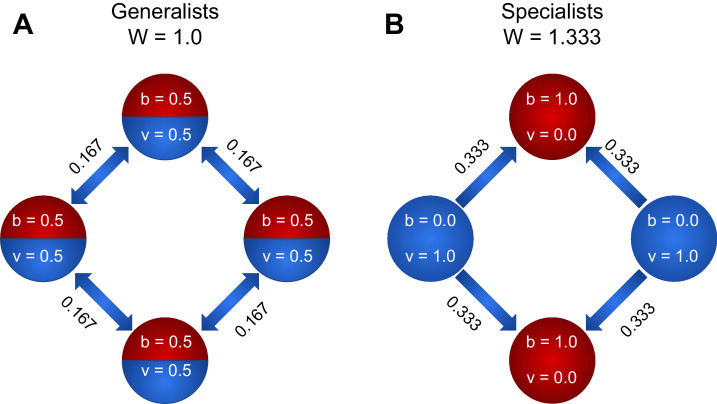
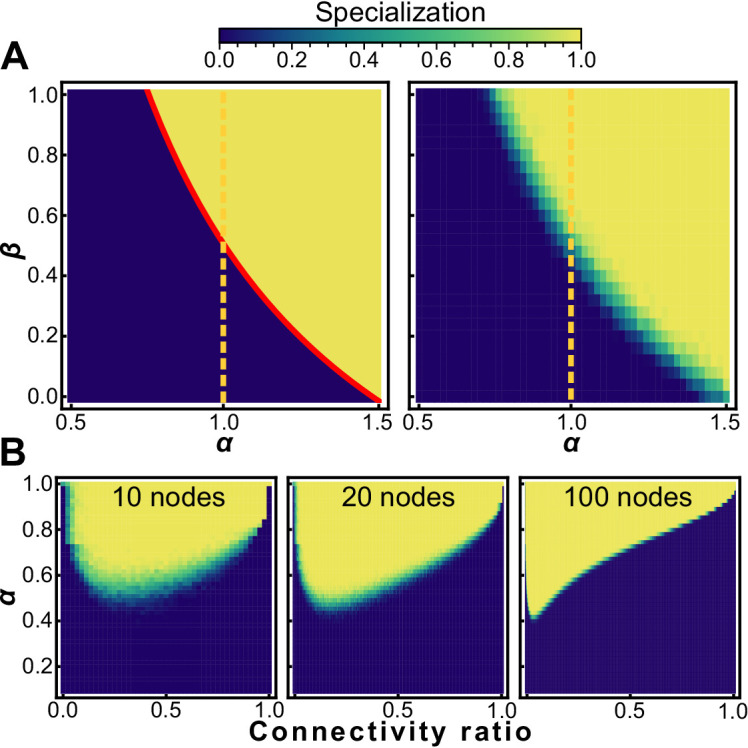
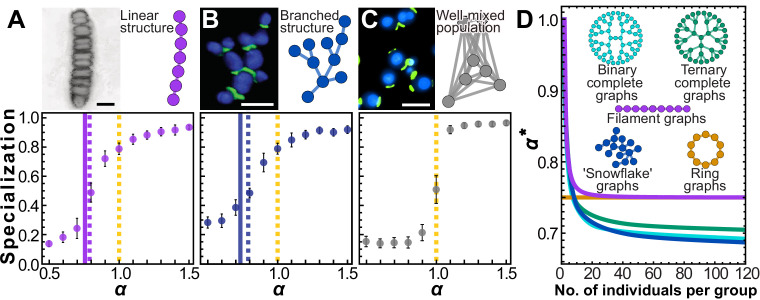
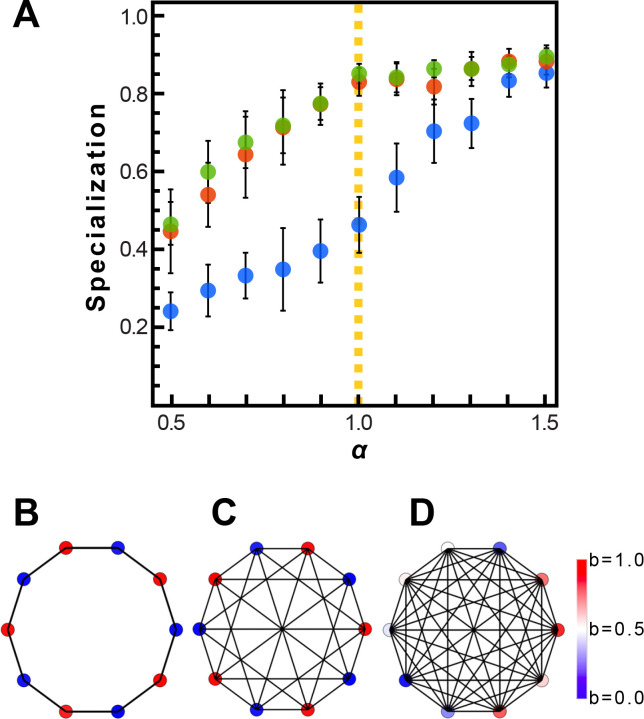
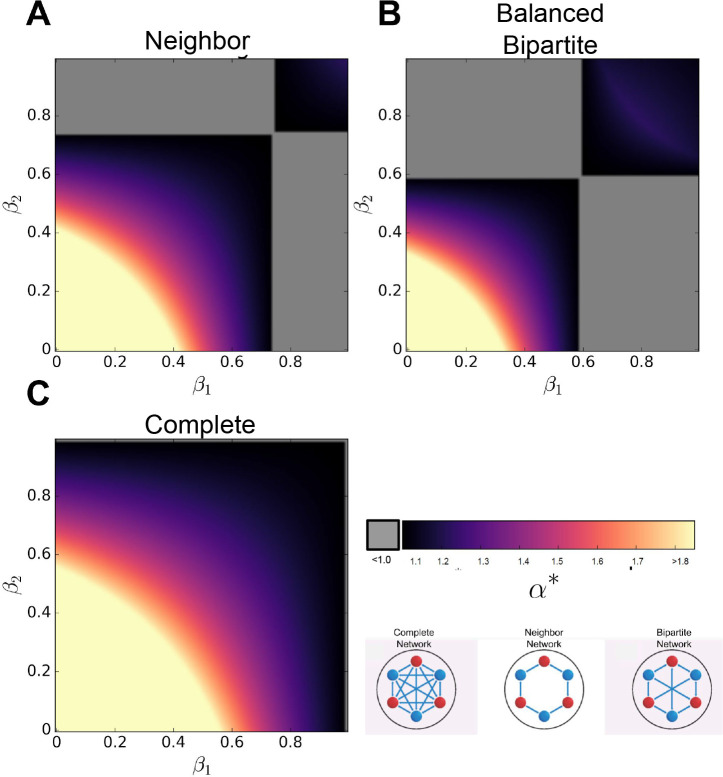
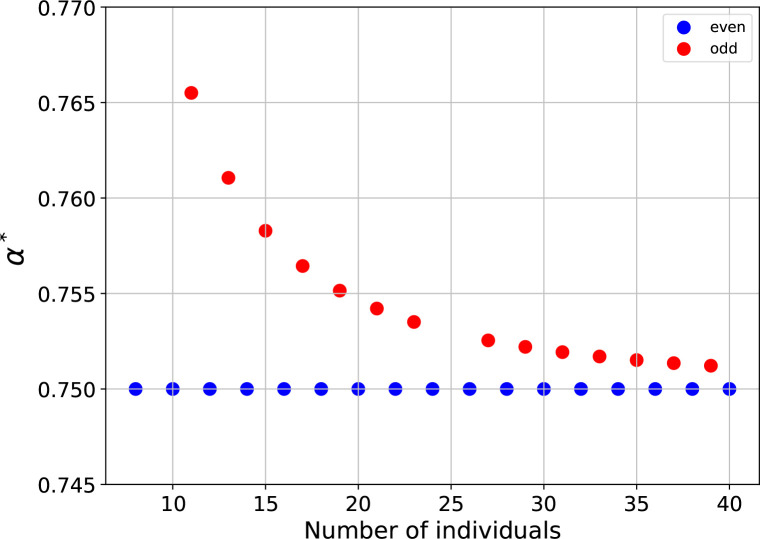
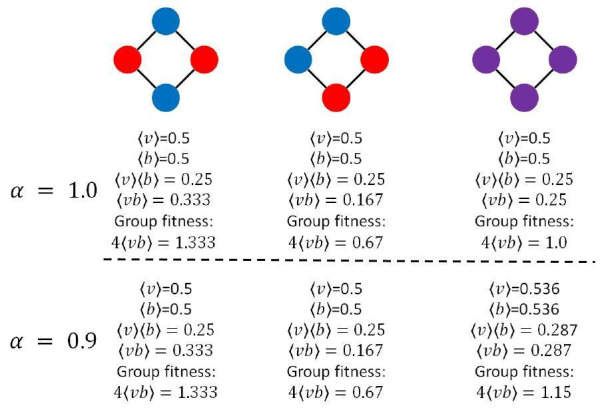
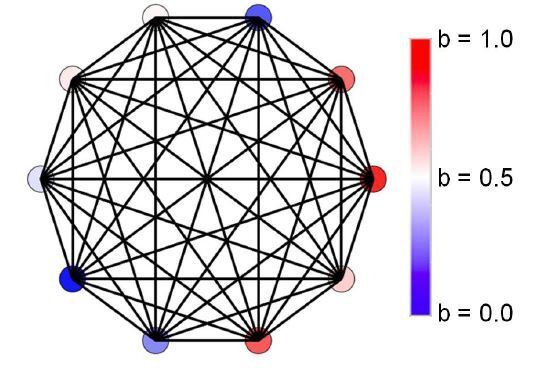
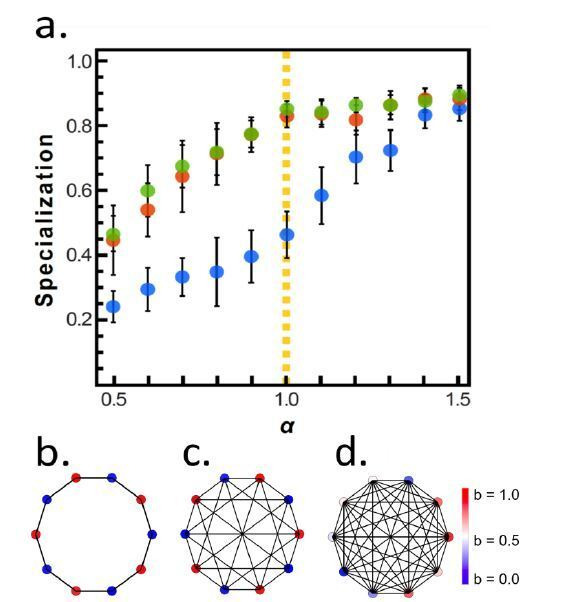
Reproductive division of labor (e.g. germ-soma specialization) is a hallmark of the evolution of multicellularity, signifying the emergence of a new type of individual and facilitating the evolution of increased organismal complexity. A large body of work from evolutionary biology, economics, and ecology has shown that specialization is beneficial when further division of labor produces an accelerating increase in absolute productivity (i.e. productivity is a convex function of specialization). Here we show that reproductive specialization is qualitatively different from classical models of resource sharing, and can evolve even when the benefits of specialization are saturating (i.e. productivity is a concave function of specialization). Through analytical theory and evolutionary individual-based simulations, we demonstrate that reproductive specialization is strongly favored in sparse networks of cellular interactions that reflect the morphology of early, simple multicellular organisms, highlighting the importance of restricted social interactions in the evolution of reproductive specialization.
Keywords: evolution; evolutionary biology; none; reproductive specialization; topology.
© 2020, Yanni et al.
Conflict of interest statement
DY, SJ, PM, JW, WR, PY No competing interests declared
Figures
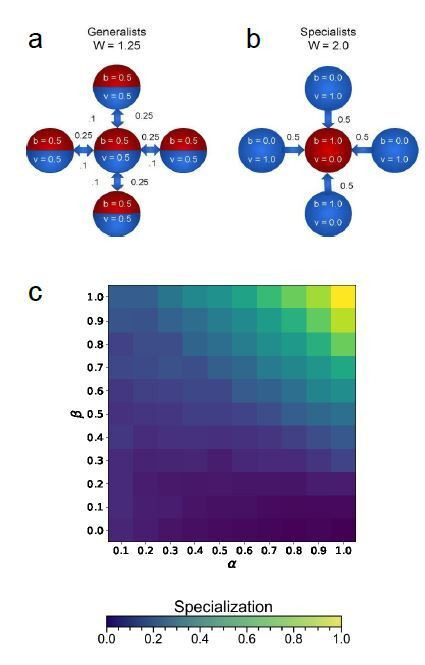
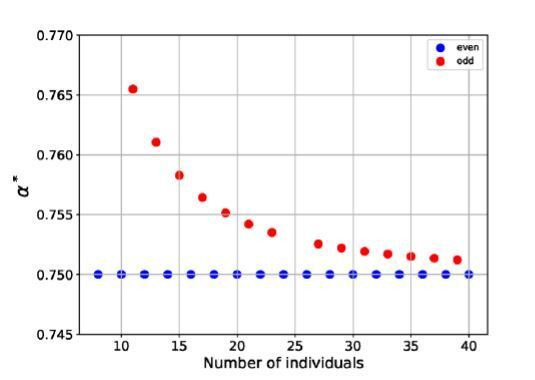
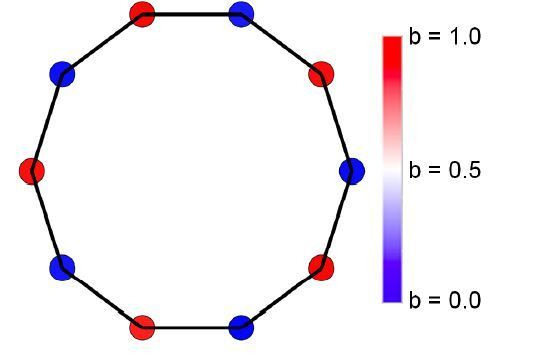
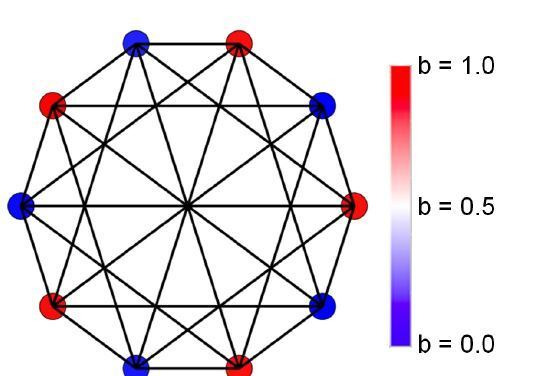
Comment in
-
How geometry shapes division of labor.Elife. 2020 Nov 3;9:e63328. doi: 10.7554/eLife.63328. Elife. 2020. PMID: 33140720 Free PMC article.
References
-
- Berman-Frank I, Quigg A, Finkel ZV, Irwin AJ, Haramaty L. Nitrogen-fixation strategies and fe requirements in cyanobacteria. Limnology and Oceanography. 2007;52:2260–2269. doi: 10.4319/lo.2007.52.5.2260. - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
