

Stressed erythrophagocytosis induces immunosuppression during sepsis through heme-mediated STAT1 dysregulation
- PMID: 32941182
- PMCID: PMC7773401
- DOI: 10.1172/JCI137468
Stressed erythrophagocytosis induces immunosuppression during sepsis through heme-mediated STAT1 dysregulation
Abstract
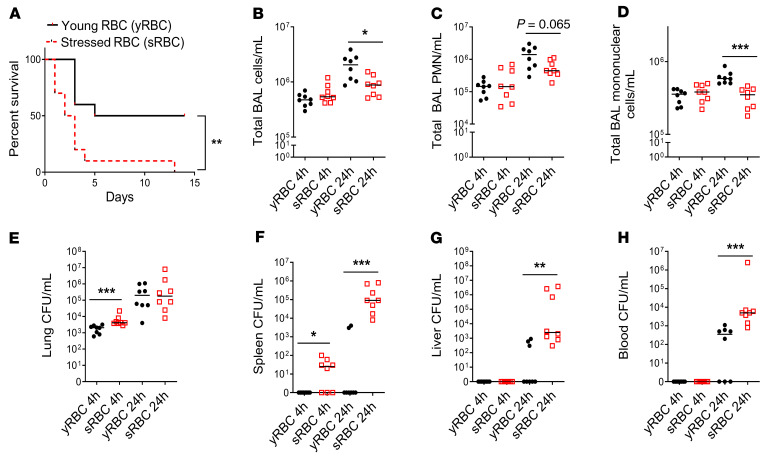
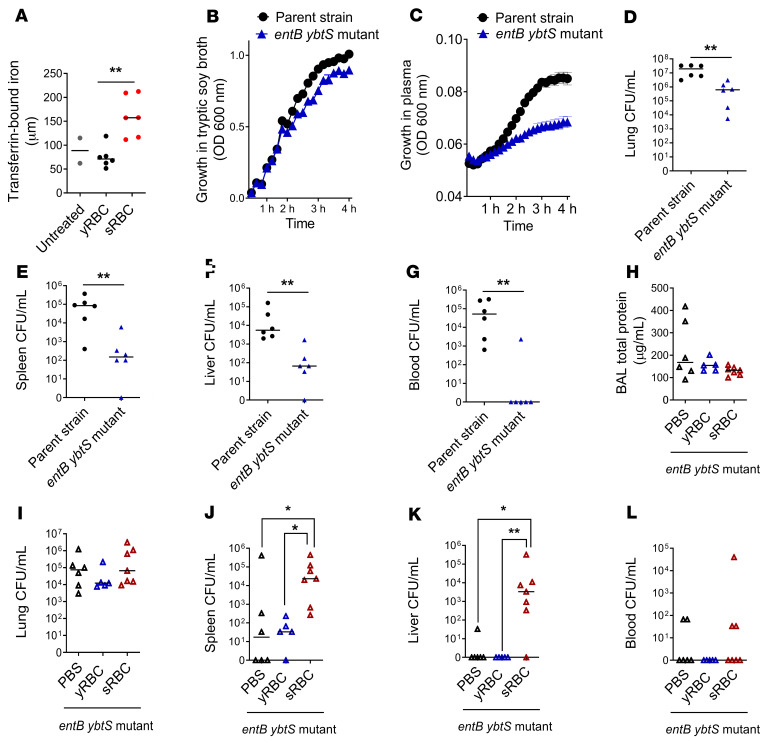
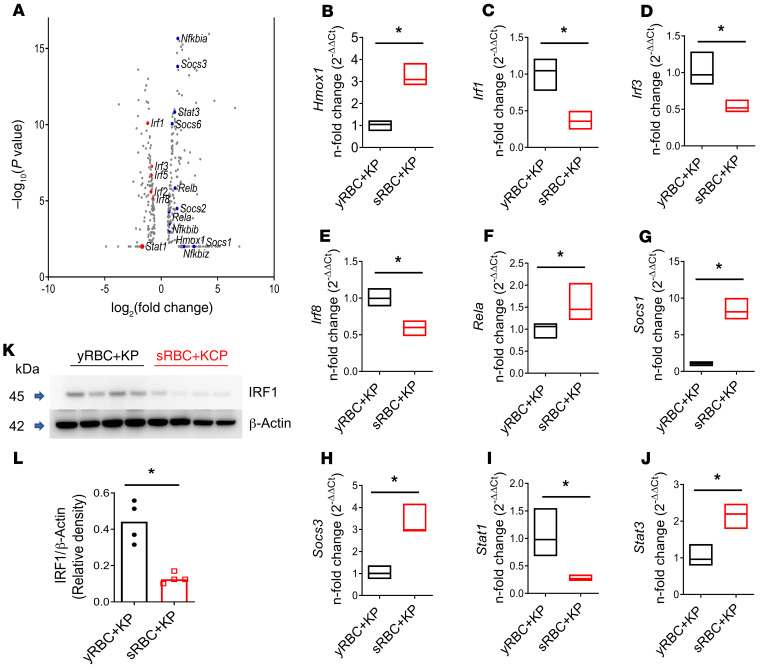
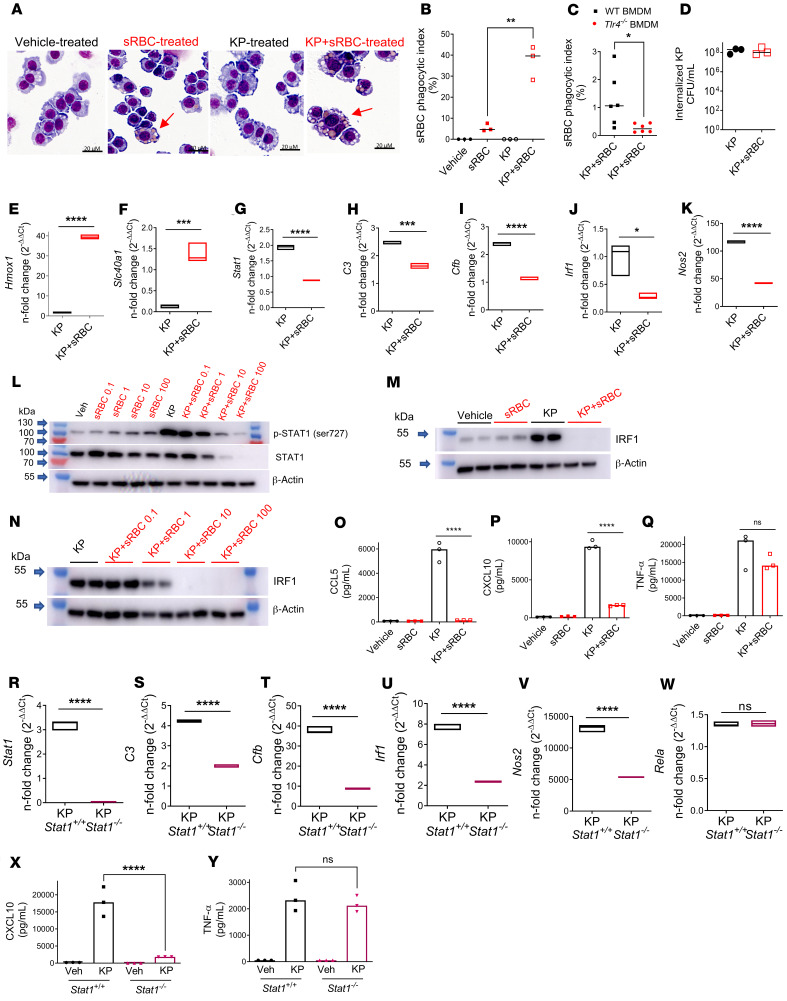
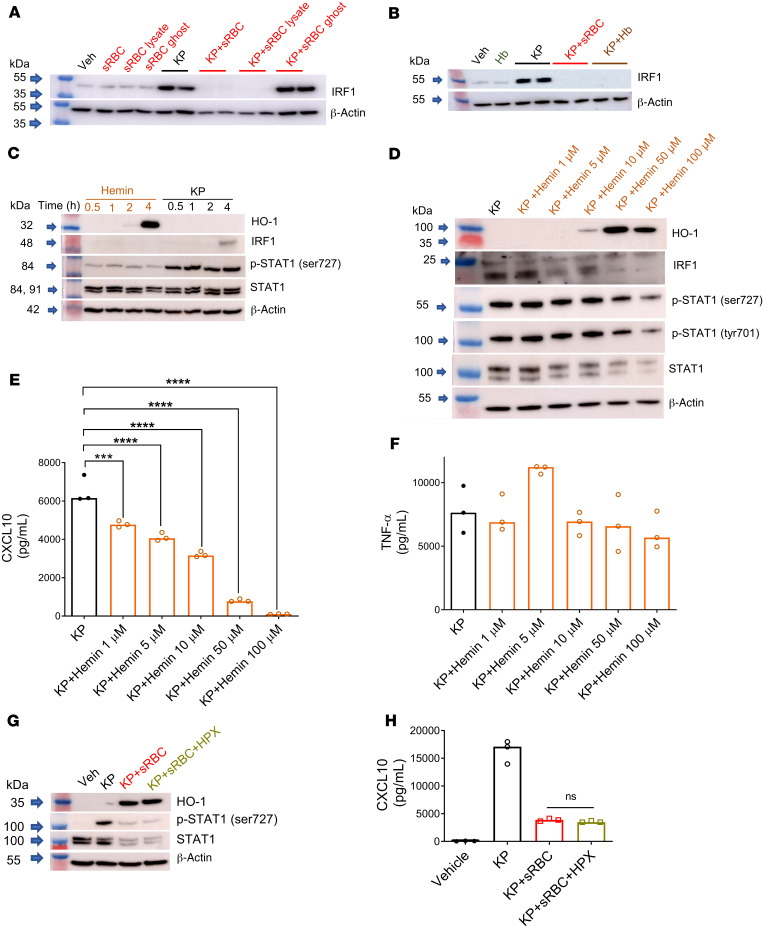
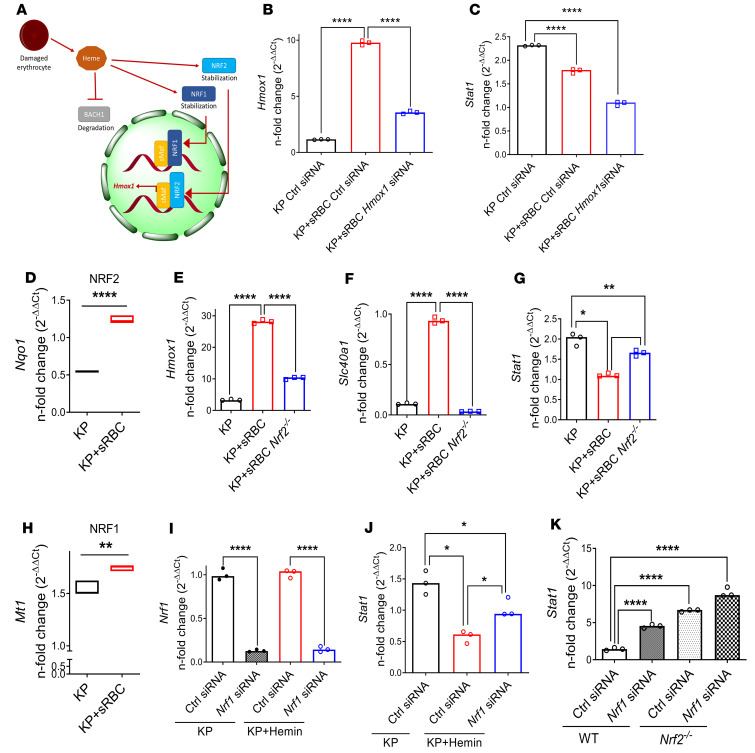
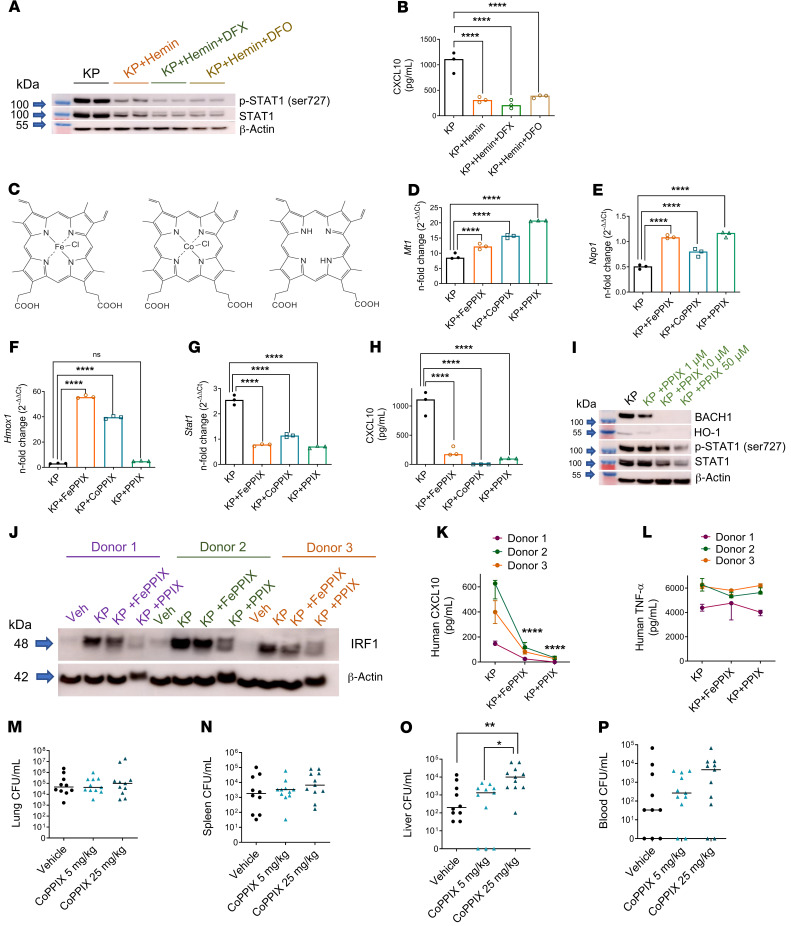
Macrophages are main effectors of heme metabolism, increasing transiently in the liver during heightened disposal of damaged or senescent RBCs (sRBCs). Macrophages are also essential in defense against microbial threats, but pathological states of heme excess may be immunosuppressive. Herein, we uncovered a mechanism whereby an acute rise in sRBC disposal by macrophages led to an immunosuppressive phenotype after intrapulmonary Klebsiella pneumoniae infection characterized by increased extrapulmonary bacterial proliferation and reduced survival from sepsis in mice. The impaired immunity to K. pneumoniae during heightened sRBC disposal was independent of iron acquisition by bacterial siderophores, in that K. pneumoniae mutants lacking siderophore function recapitulated the findings observed with the WT strain. Rather, sRBC disposal induced a liver transcriptomic profile notable for suppression of Stat1 and IFN-related responses during K. pneumoniae sepsis. Excess heme handling by macrophages recapitulated STAT1 suppression during infection that required synergistic NRF1 and NRF2 activation but was independent of heme oxygenase-1 induction. Whereas iron was dispensable, the porphyrin moiety of heme was sufficient to mediate suppression of STAT1-dependent responses in human and mouse macrophages and promoted liver dissemination of K. pneumoniae in vivo. Thus, cellular heme metabolism dysfunction negatively regulated the STAT1 pathway, with implications in severe infection.
Keywords: Bacterial infections; Infectious disease; Innate immunity; Macrophages; Pulmonology.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
