

Eukaryotic cell biology is temporally coordinated to support the energetic demands of protein homeostasis
- PMID: 32943618
- PMCID: PMC7499178
- DOI: 10.1038/s41467-020-18330-x
Eukaryotic cell biology is temporally coordinated to support the energetic demands of protein homeostasis
Erratum in
-
Author Correction: Eukaryotic cell biology is temporally coordinated to support the energetic demands of protein homeostasis.Nat Commun. 2021 Dec 8;12(1):7269. doi: 10.1038/s41467-021-27497-w. Nat Commun. 2021. PMID: 34880246 Free PMC article. No abstract available.
Abstract
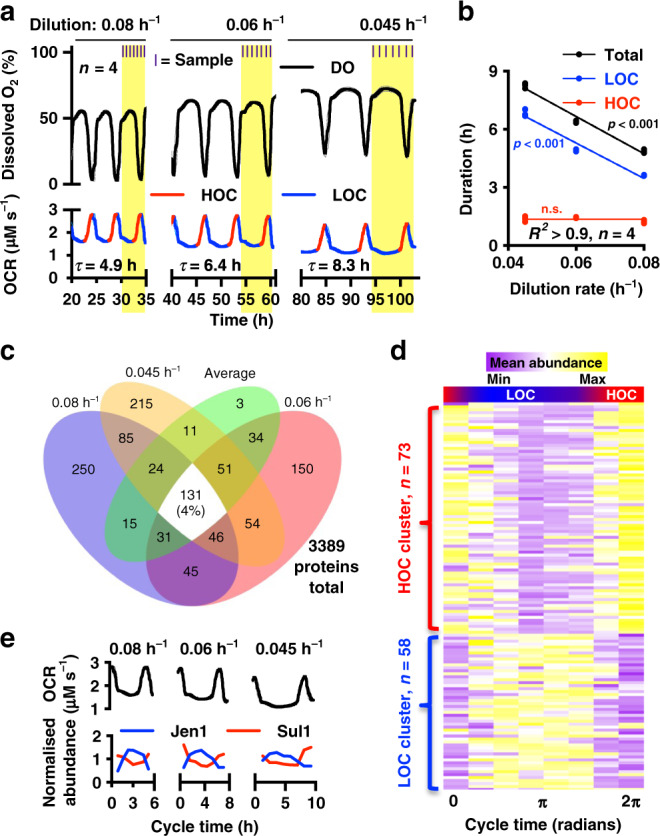
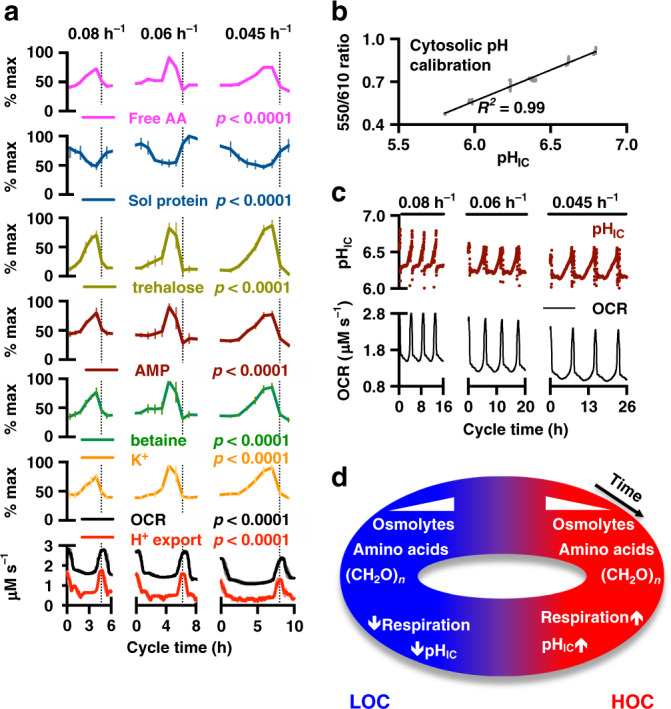
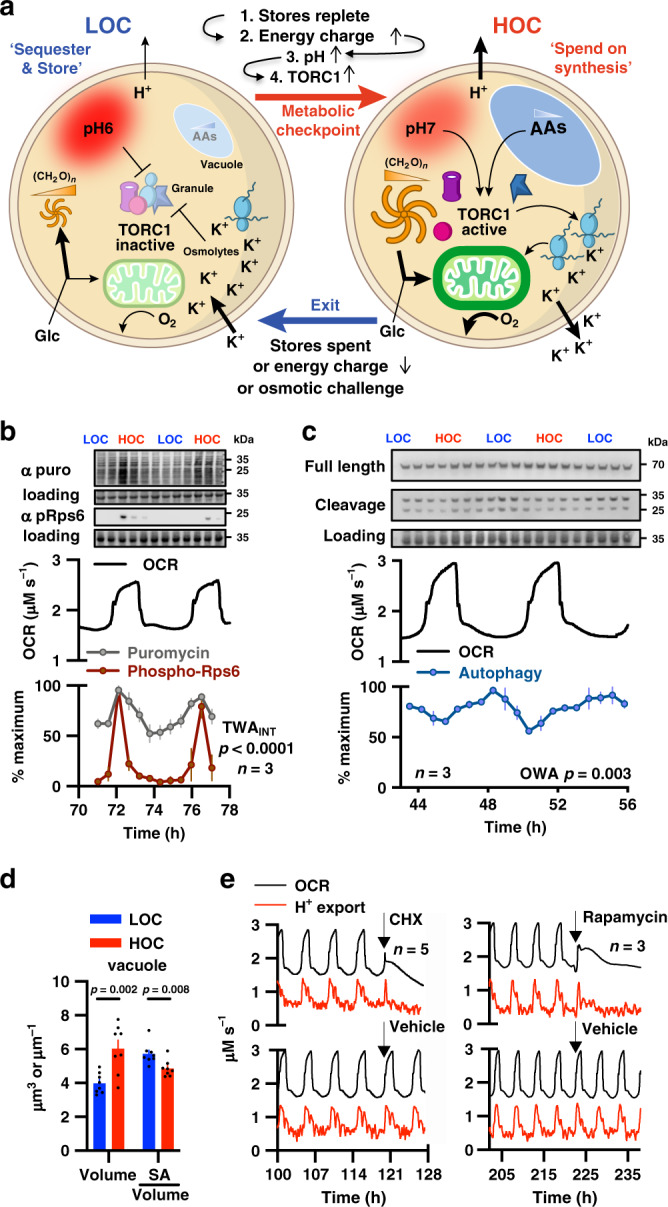
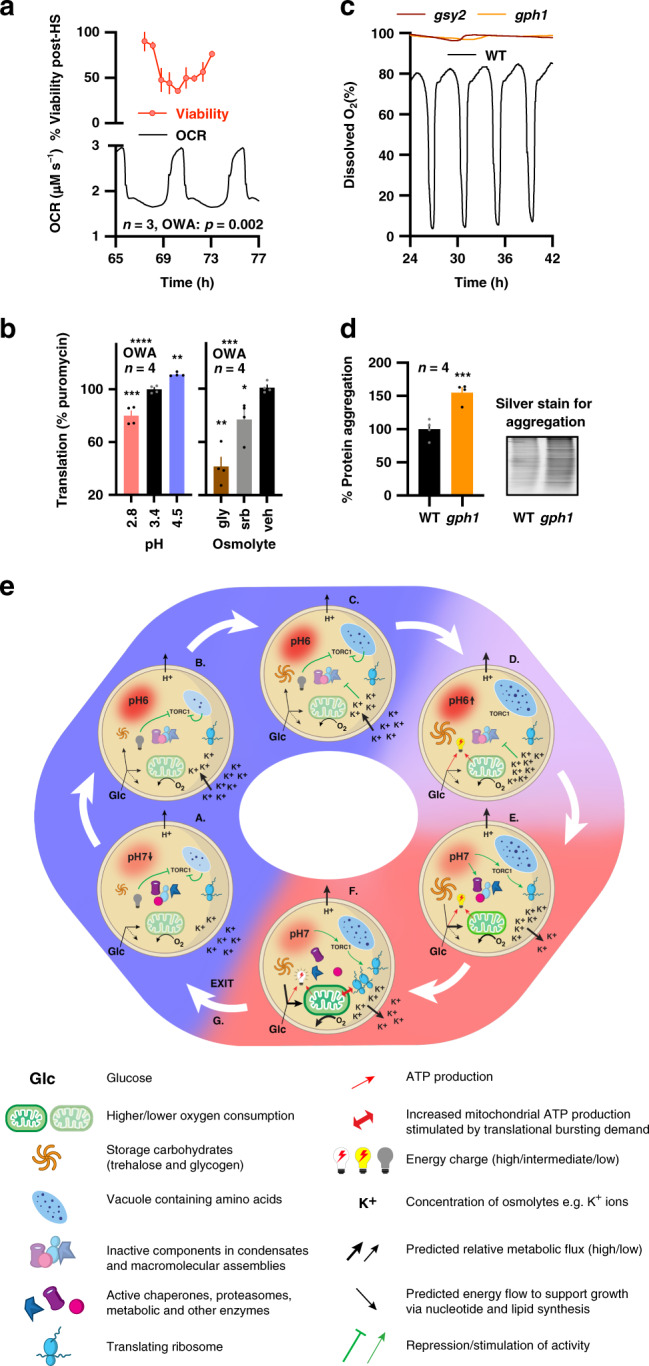
Yeast physiology is temporally regulated, this becomes apparent under nutrient-limited conditions and results in respiratory oscillations (YROs). YROs share features with circadian rhythms and interact with, but are independent of, the cell division cycle. Here, we show that YROs minimise energy expenditure by restricting protein synthesis until sufficient resources are stored, while maintaining osmotic homeostasis and protein quality control. Although nutrient supply is constant, cells sequester and store metabolic resources via increased transport, autophagy and biomolecular condensation. Replete stores trigger increased H+ export which stimulates TORC1 and liberates proteasomes, ribosomes, chaperones and metabolic enzymes from non-membrane bound compartments. This facilitates translational bursting, liquidation of storage carbohydrates, increased ATP turnover, and the export of osmolytes. We propose that dynamic regulation of ion transport and metabolic plasticity are required to maintain osmotic and protein homeostasis during remodelling of eukaryotic proteomes, and that bioenergetic constraints selected for temporal organisation that promotes oscillatory behaviour.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Lloyd D. Respiratory oscillations in yeasts. Adv. Exp. Med. Biol. 2008;641:118–140. - PubMed
-
- Cai L, Tu BP. Driving the cell cycle through metabolism. Annu Rev. Cell Dev. Biol. 2012;28:59–87. - PubMed
-
- Beuse M, Bartling R, Kopmann A, Diekmann H, Thoma M. Effect of the dilution rate on the mode of oscillation in continuous cultures of Saccharomyces cerevisiae. J. Biotechnol. 1998;61:15–31. - PubMed
-
- Causton, H. C. Metabolic Rhythms: a framework for coordinating cellular function. Eur. J. Neurosci., 10.1111/ejn.14296 (2018). - PubMed
-
- Mellor J. The molecular basis of metabolic cycles and their relationship to circadian rhythms. Nat. Struct. Mol. Biol. 2016;23:1035–1044. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
