Locus coeruleus: a new look at the blue spot
- PMID: 32943779
- PMCID: PMC8991985
- DOI: 10.1038/s41583-020-0360-9
Locus coeruleus: a new look at the blue spot
Abstract
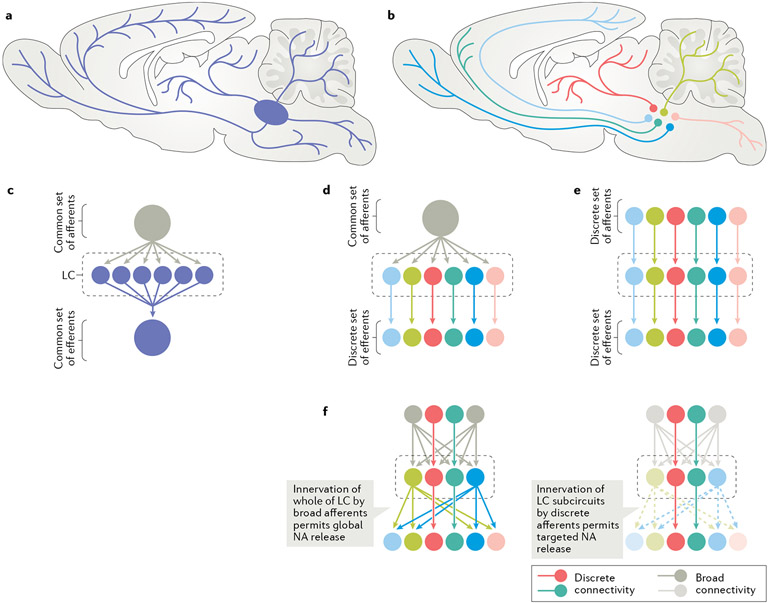
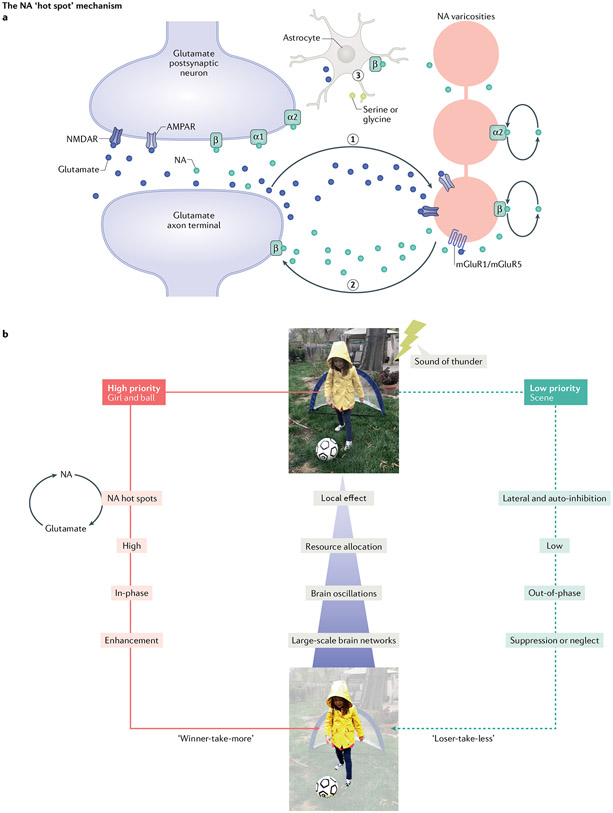
The locus coeruleus (LC), or 'blue spot', is a small nucleus located deep in the brainstem that provides the far-reaching noradrenergic neurotransmitter system of the brain. This phylogenetically conserved nucleus has proved relatively intractable to full characterization, despite more than 60 years of concerted efforts by investigators. Recently, an array of powerful new neuroscience tools have provided unprecedented access to this elusive nucleus, revealing new levels of organization and function. We are currently at the threshold of major discoveries regarding how this tiny brainstem structure exerts such varied and significant influences over brain function and behaviour. All LC neurons receive inputs related to autonomic arousal, but distinct subpopulations of those neurons can encode specific cognitive processes, presumably through more specific inputs from the forebrain areas. This ability, combined with specific patterns of innervation of target areas and heterogeneity in receptor distributions, suggests that activation of the LC has more specific influences on target networks than had initially been imagined.
Figures