

pH-Gated Succinate Secretion Regulates Muscle Remodeling in Response to Exercise
- PMID: 32946811
- PMCID: PMC7778787
- DOI: 10.1016/j.cell.2020.08.039
pH-Gated Succinate Secretion Regulates Muscle Remodeling in Response to Exercise
Abstract
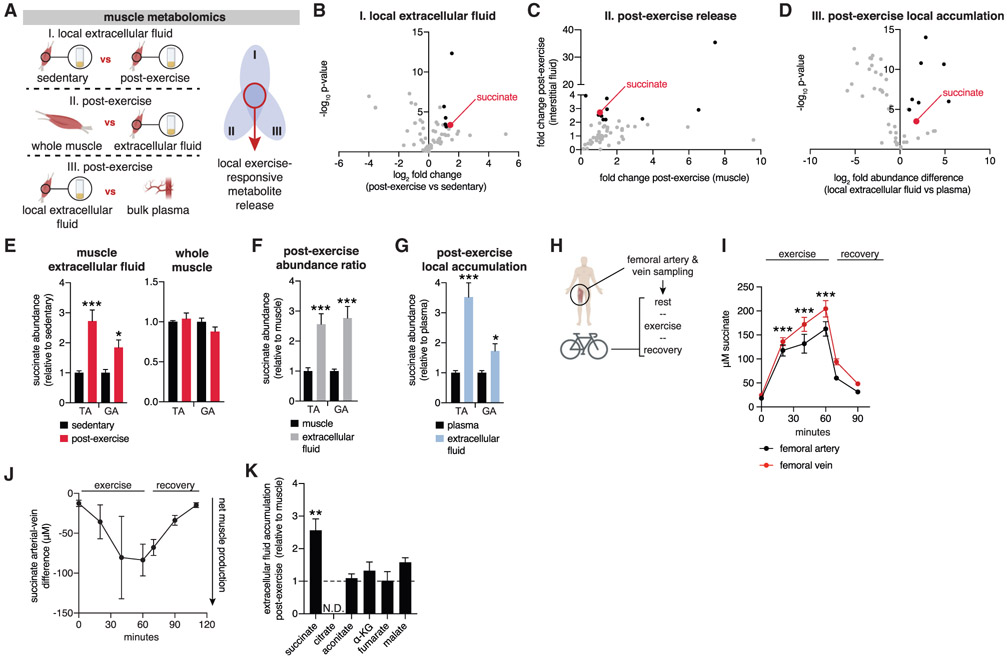
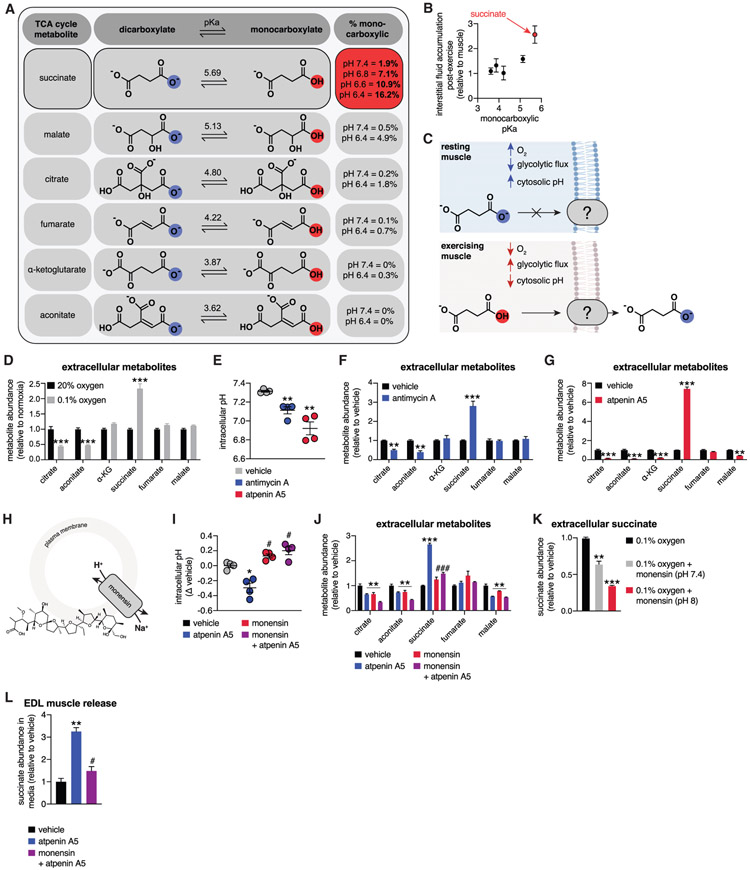
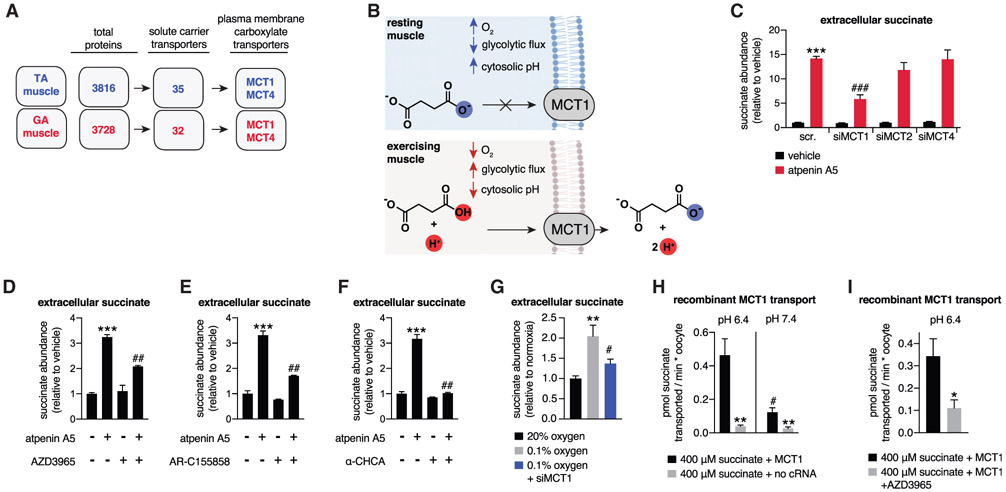
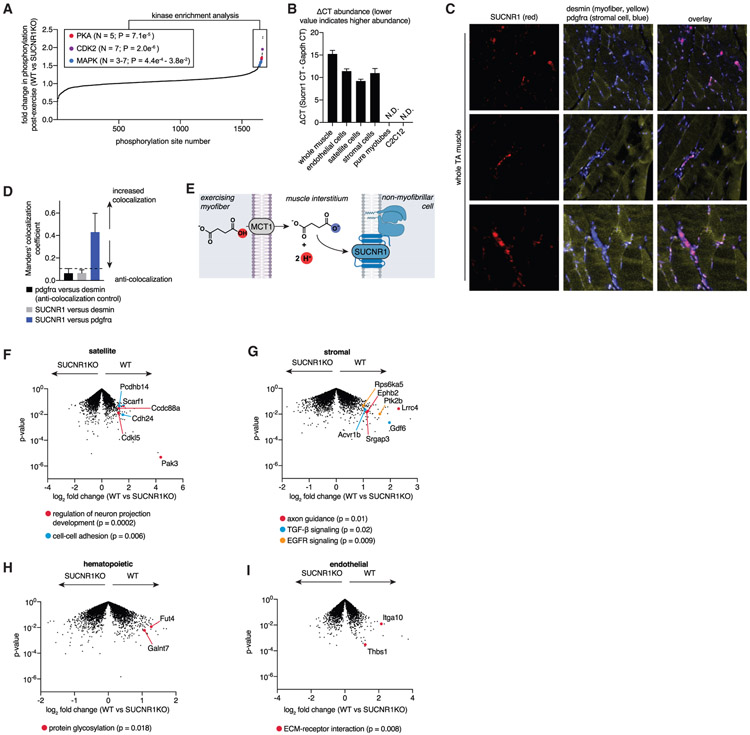
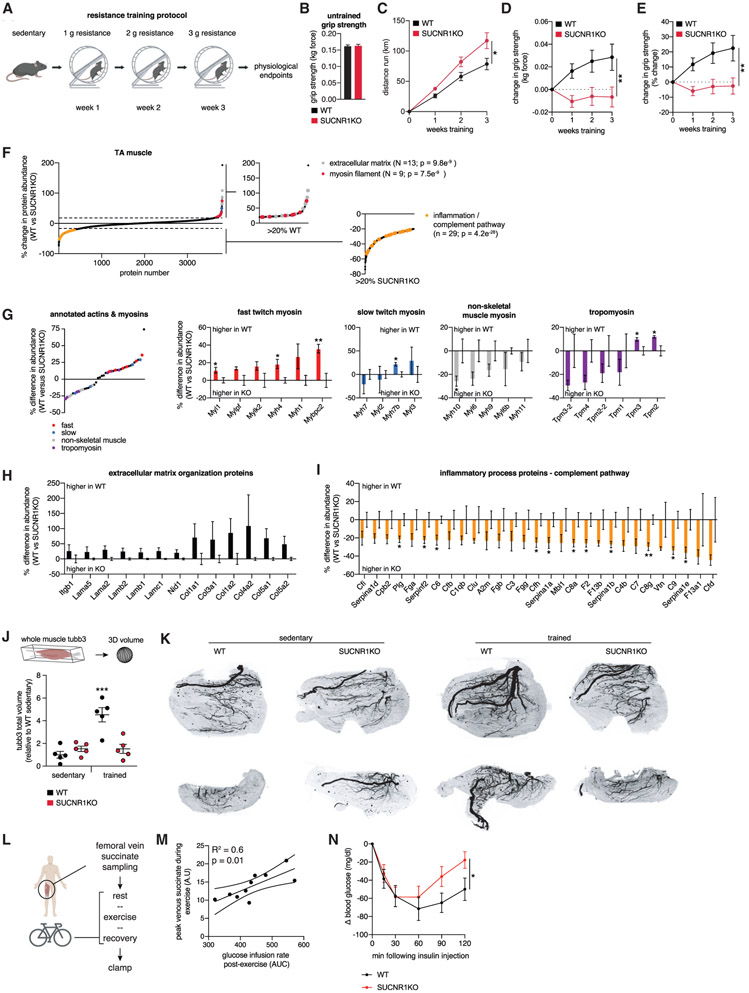
In response to skeletal muscle contraction during exercise, paracrine factors coordinate tissue remodeling, which underlies this healthy adaptation. Here we describe a pH-sensing metabolite signal that initiates muscle remodeling upon exercise. In mice and humans, exercising skeletal muscle releases the mitochondrial metabolite succinate into the local interstitium and circulation. Selective secretion of succinate is facilitated by its transient protonation, which occurs upon muscle cell acidification. In the protonated monocarboxylic form, succinate is rendered a transport substrate for monocarboxylate transporter 1, which facilitates pH-gated release. Upon secretion, succinate signals via its cognate receptor SUCNR1 in non-myofibrillar cells in muscle tissue to control muscle-remodeling transcriptional programs. This succinate-SUCNR1 signaling is required for paracrine regulation of muscle innervation, muscle matrix remodeling, and muscle strength in response to exercise training. In sum, we define a bioenergetic sensor in muscle that utilizes intracellular pH and succinate to coordinate tissue adaptation to exercise.
Keywords: SUCNR1; exercise; innervation; muscle; succinate.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests E.T.C. has filed for a patent based on data describing the role of SUCNR1 agonism in regulation of muscle remodeling in this work.
Figures
Comment in
-
Succinate regulates muscle exercise adaptations.Nat Rev Endocrinol. 2020 Dec;16(12):678-679. doi: 10.1038/s41574-020-00429-2. Nat Rev Endocrinol. 2020. PMID: 33009503 No abstract available.
References
-
- Barker D, and Saito M (1981). Autonomic innervation of receptors and muscle fibres in cat skeletal muscle. Proc. R. Soc. Lond. B Biol. Sci 212,317–332. - PubMed
-
- Beitzel F, Gregorevic P, Ryall JG, Plant DR, Sillence MN, and Lynch GS (2004). Beta2-adrenoceptor agonist fenoterol enhances functional repair of regenerating rat skeletal muscle after injury. J. Appl. Physiol 96, 1385–1392. - PubMed
-
- Bright CM, and Ellis D (1992). Intracellular pH changes induced by hypoxia and anoxia in isolated sheep heart Purkinje fibres. Exp. Physiol 77, 165–175. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
