

V(DD)J recombination is an important and evolutionarily conserved mechanism for generating antibodies with unusually long CDR3s
- PMID: 32948615
- PMCID: PMC7605257
- DOI: 10.1101/gr.259598.119
V(DD)J recombination is an important and evolutionarily conserved mechanism for generating antibodies with unusually long CDR3s
Abstract
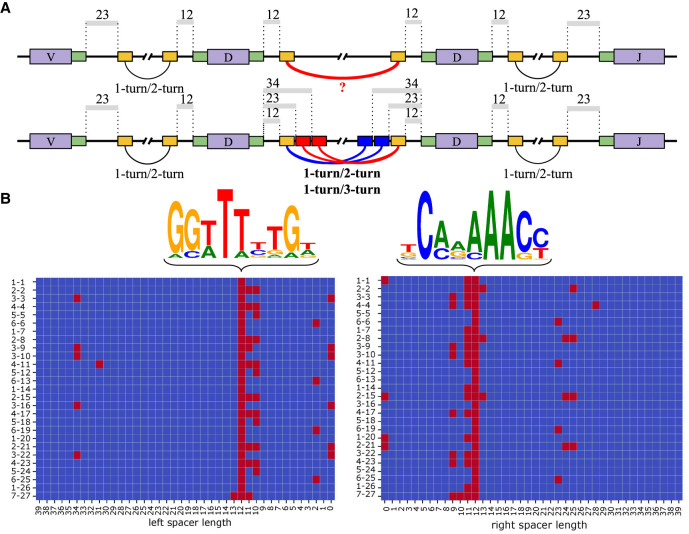
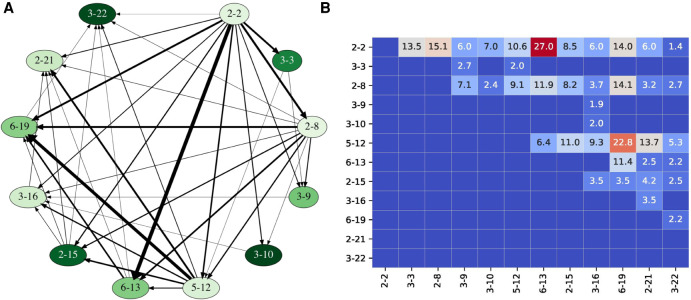
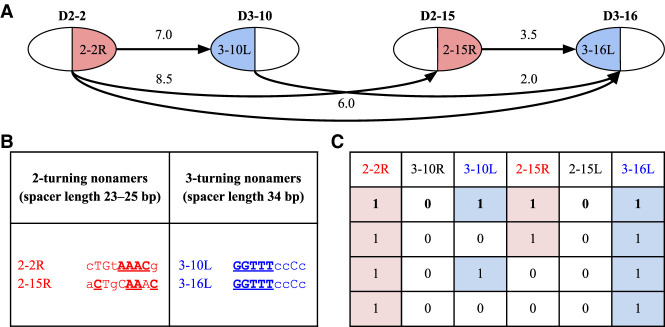
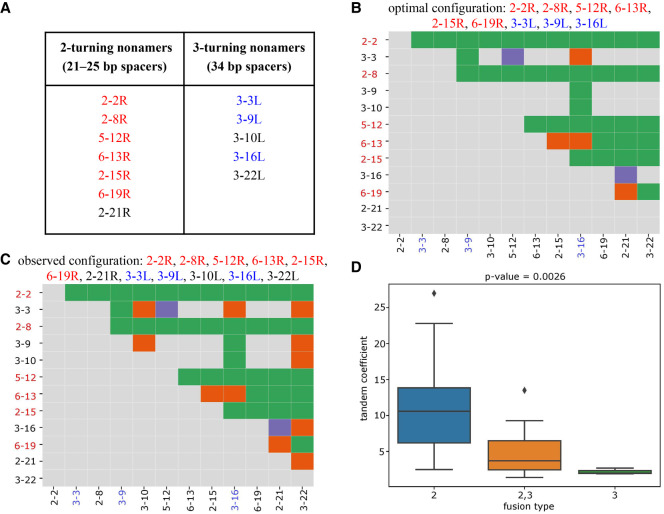
The V(DD)J recombination is currently viewed as an aberrant and inconsequential variant of the canonical V(D)J recombination. Moreover, since the classical 12/23 rule for the V(D)J recombination fails to explain the V(DD)J recombination, the molecular mechanism of tandem D-D fusions has remained unknown since they were discovered three decades ago. Revealing this mechanism is a biomedically important goal since tandem fusions contribute to broadly neutralizing antibodies with ultralong CDR3s. We reveal previously overlooked cryptic nonamers in the recombination signal sequences of human IGHD genes and demonstrate that these nonamers explain the vast majority of tandem fusions in human repertoires. We further reveal large clonal lineages formed by tandem fusions in antigen-stimulated immunosequencing data sets, suggesting that such data sets contain many more tandem fusions than previously thought and that about a quarter of large clonal lineages with unusually long CDR3s are generated through tandem fusions. Finally, we developed the SEARCH-D algorithm for identifying D genes in mammalian genomes and applied it to the recently completed Vertebrate Genomes Project assemblies, nearly doubling the number of mammalian species with known D genes. Our analysis revealed cryptic nonamers in RSSs of many mammalian genomes, thus demonstrating that the V(DD)J recombination is not a "bug" but an important feature preserved throughout mammalian evolution.
© 2020 Safonova and Pevzner; Published by Cold Spring Harbor Laboratory Press.
Figures

Similar articles
-
De novo Inference of Diversity Genes and Analysis of Non-canonical V(DD)J Recombination in Immunoglobulins.Front Immunol. 2019 May 3;10:987. doi: 10.3389/fimmu.2019.00987. eCollection 2019. Front Immunol. 2019. PMID: 31134072 Free PMC article.
-
Frequency and genetic characterization of V(DD)J recombinants in the human peripheral blood antibody repertoire.Immunology. 2012 Sep;137(1):56-64. doi: 10.1111/j.1365-2567.2012.03605.x. Immunology. 2012. PMID: 22612413 Free PMC article.
-
HTS and scRNA-seq revealed that the location and RSS quality of the mammalian TRBV and TRBJ genes impact biased rearrangement.BMC Genomics. 2024 Oct 29;25(1):1010. doi: 10.1186/s12864-024-10887-x. BMC Genomics. 2024. PMID: 39472808 Free PMC article.
-
Diversity in the Cow Ultralong CDR H3 Antibody Repertoire.Front Immunol. 2018 Jun 4;9:1262. doi: 10.3389/fimmu.2018.01262. eCollection 2018. Front Immunol. 2018. PMID: 29915599 Free PMC article. Review.
-
V(D)J recombination, somatic hypermutation and class switch recombination of immunoglobulins: mechanism and regulation.Immunology. 2020 Jul;160(3):233-247. doi: 10.1111/imm.13176. Epub 2020 Feb 27. Immunology. 2020. PMID: 32031242 Free PMC article. Review.
Cited by
-
Functional antibodies exhibit light chain coherence.Nature. 2022 Nov;611(7935):352-357. doi: 10.1038/s41586-022-05371-z. Epub 2022 Oct 26. Nature. 2022. PMID: 36289331 Free PMC article.
-
Unveiling inverted D genes and D-D fusions in human antibody repertoires unlocks novel antibody diversity.Commun Biol. 2025 Jan 28;8(1):133. doi: 10.1038/s42003-024-07441-6. Commun Biol. 2025. PMID: 39875530 Free PMC article.
-
Novel bimodal TRBD1-TRBD2 rearrangements with dual or absent D-region contribute to TRB V-(D)-J combinatorial diversity.Front Immunol. 2023 Sep 7;14:1245175. doi: 10.3389/fimmu.2023.1245175. eCollection 2023. Front Immunol. 2023. PMID: 37744336 Free PMC article.
-
UniAligner: a parameter-free framework for fast sequence alignment.Nat Methods. 2023 Sep;20(9):1346-1354. doi: 10.1038/s41592-023-01970-4. Epub 2023 Aug 14. Nat Methods. 2023. PMID: 37580559
-
ALIGaToR: A tool for the automated annotation of immunoglobulin and T cell receptor genomic loci.bioRxiv [Preprint]. 2025 May 8:2025.05.02.651960. doi: 10.1101/2025.05.02.651960. bioRxiv. 2025. PMID: 40568063 Free PMC article. Preprint.
References
-
- Corbett SJ, Tomlinson IM, Sonnhammer EL, Buck D, Winter G. 1997. Sequence of the human immunoglobulin diversity (D) segment locus: A systematic analysis provides no evidence for the use of DIR segments, inverted D segments, “minor” D segments or D-D recombination. J Mol Biol 270: 587–597. 10.1006/jmbi.1997.1141 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources