

Transcription factor expression defines subclasses of developing projection neurons highly similar to single-cell RNA-seq subtypes
- PMID: 32948690
- PMCID: PMC7547209
- DOI: 10.1073/pnas.2008013117
Transcription factor expression defines subclasses of developing projection neurons highly similar to single-cell RNA-seq subtypes
Abstract
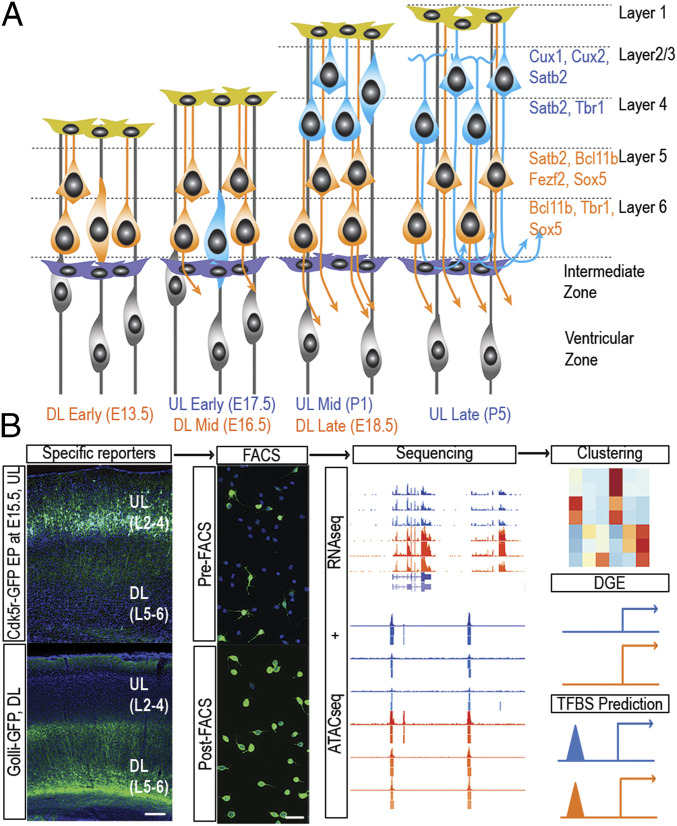
We are only just beginning to catalog the vast diversity of cell types in the cerebral cortex. Such categorization is a first step toward understanding how diversification relates to function. All cortical projection neurons arise from a uniform pool of progenitor cells that lines the ventricles of the forebrain. It is still unclear how these progenitor cells generate the more than 50 unique types of mature cortical projection neurons defined by their distinct gene-expression profiles. Moreover, exactly how and when neurons diversify their function during development is unknown. Here we relate gene expression and chromatin accessibility of two subclasses of projection neurons with divergent morphological and functional features as they develop in the mouse brain between embryonic day 13 and postnatal day 5 in order to identify transcriptional networks that diversify neuron cell fate. We compare these gene-expression profiles with published profiles of single cells isolated from similar populations and establish that layer-defined cell classes encompass cell subtypes and developmental trajectories identified using single-cell sequencing. Given the depth of our sequencing, we identify groups of transcription factors with particularly dense subclass-specific regulation and subclass-enriched transcription factor binding motifs. We also describe transcription factor-adjacent long noncoding RNAs that define each subclass and validate the function of Myt1l in balancing the ratio of the two subclasses in vitro. Our multidimensional approach supports an evolving model of progressive restriction of cell fate competence through inherited transcriptional identities.
Keywords: cortical development; gene regulation; next-generation sequencing; transcription.
Conflict of interest statement
The authors declare no competing interest.
Figures





Similar articles
-
Prospective separation and transcriptome analyses of cortical projection neurons and interneurons based on lineage tracing by Tbr2 (Eomes)-GFP/Dcx-mRFP reporters.Dev Neurobiol. 2016 Jun;76(6):587-99. doi: 10.1002/dneu.22332. Epub 2015 Aug 17. Dev Neurobiol. 2016. PMID: 26248544 Free PMC article.
-
Fezf2 expression in layer 5 projection neurons of mature mouse motor cortex.J Comp Neurol. 2016 Mar 1;524(4):829-45. doi: 10.1002/cne.23875. Epub 2015 Aug 30. J Comp Neurol. 2016. PMID: 26234885
-
Transcription factor programming of human ES cells generates functional neurons expressing both upper and deep layer cortical markers.PLoS One. 2018 Oct 11;13(10):e0204688. doi: 10.1371/journal.pone.0204688. eCollection 2018. PLoS One. 2018. PMID: 30307948 Free PMC article.
-
From radial glia to pyramidal-projection neuron: transcription factor cascades in cerebral cortex development.Mol Neurobiol. 2006 Feb;33(1):33-50. doi: 10.1385/MN:33:1:033. Mol Neurobiol. 2006. PMID: 16388109 Review.
-
The determination of projection neuron identity in the developing cerebral cortex.Curr Opin Neurobiol. 2008 Feb;18(1):28-35. doi: 10.1016/j.conb.2008.05.006. Epub 2008 May 26. Curr Opin Neurobiol. 2008. PMID: 18508260 Free PMC article. Review.
Cited by
-
New Molecular Players in the Development of Callosal Projections.Cells. 2020 Dec 26;10(1):29. doi: 10.3390/cells10010029. Cells. 2020. PMID: 33375263 Free PMC article. Review.
-
Forebrain Neural Precursor Cells Are Differentially Vulnerable to Zika Virus Infection.eNeuro. 2021 Sep 9;8(5):ENEURO.0108-21.2021. doi: 10.1523/ENEURO.0108-21.2021. Print 2021 Sep-Oct. eNeuro. 2021. PMID: 34272257 Free PMC article.
-
SATB2-LEMD2 interaction links nuclear shape plasticity to regulation of cognition-related genes.EMBO J. 2021 Feb 1;40(3):e103701. doi: 10.15252/embj.2019103701. Epub 2020 Dec 15. EMBO J. 2021. PMID: 33319920 Free PMC article.
-
Single-cell analysis reveals specific neuronal transition during mouse corticogenesis.Front Cell Dev Biol. 2023 Nov 6;11:1209320. doi: 10.3389/fcell.2023.1209320. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 38020907 Free PMC article.
-
Diverse axonal morphologies of individual callosal projection neurons reveal new insights into brain connectivity.Curr Opin Neurobiol. 2024 Feb;84:102837. doi: 10.1016/j.conb.2023.102837. Epub 2024 Jan 24. Curr Opin Neurobiol. 2024. PMID: 38271848 Free PMC article. Review.
References
-
- Zeng H., Sanes J. R., Neuronal cell-type classification: Challenges, opportunities and the path forward. Nat. Rev. Neurosci. 18, 530–546 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
