

GM2 ganglioside accumulation causes neuroinflammation and behavioral alterations in a mouse model of early onset Tay-Sachs disease
- PMID: 32951593
- PMCID: PMC7504627
- DOI: 10.1186/s12974-020-01947-6
GM2 ganglioside accumulation causes neuroinflammation and behavioral alterations in a mouse model of early onset Tay-Sachs disease
Abstract
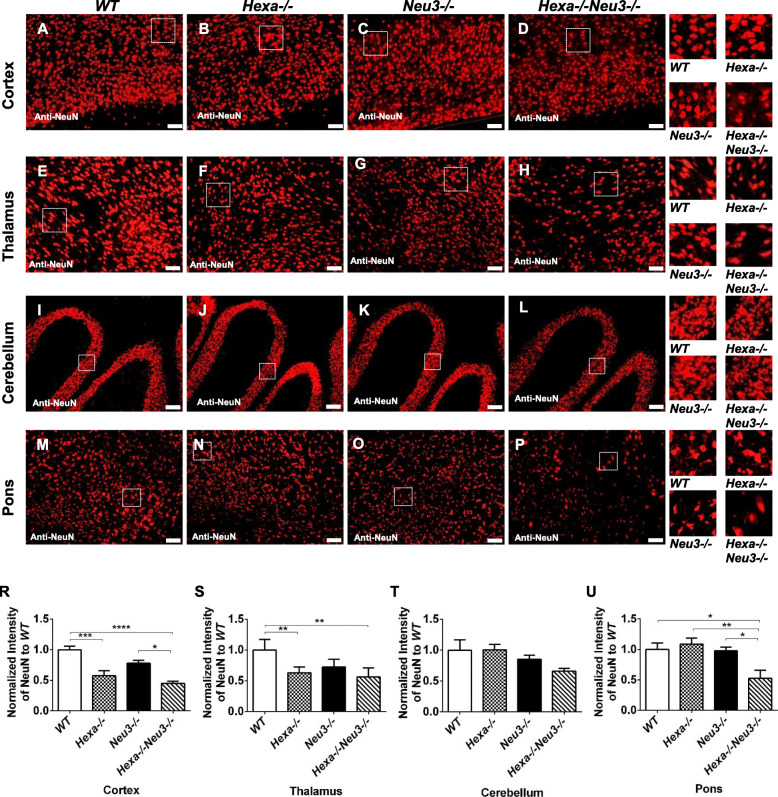
Background: Tay-Sachs disease (TSD), a type of GM2-gangliosidosis, is a progressive neurodegenerative lysosomal storage disorder caused by mutations in the α subunit of the lysosomal β-hexosaminidase enzyme. This disease is characterized by excessive accumulation of GM2 ganglioside, predominantly in the central nervous system. Although Tay-Sachs patients appear normal at birth, the progressive accumulation of undegraded GM2 gangliosides in neurons leads to death. Recently, an early onset Tay-Sachs disease mouse model, with genotype Hexa-/-Neu3-/-, was generated. Progressive accumulation of GM2 led to premature death of the double KO mice. Importantly, this double-deficient mouse model displays typical features of Tay-Sachs patients, such as cytoplasmic vacuolization of nerve cells, deterioration of Purkinje cells, neuronal death, deceleration in movement, ataxia, and tremors. GM2-gangliosidosis is characterized by acute neurodegeneration preceded by activated microglia expansion, macrophage, and astrocyte activation, along with the production of inflammatory mediators. However, the mechanism of disease progression in Hexa-/-Neu3-/- mice, relevant to neuroinflammation is poorly understood.
Method: In this study, we investigated the onset and progression of neuroinflammatory changes in the cortex, cerebellum, and retina of Hexa-/-Neu3-/- mice and control littermates by using a combination of molecular genetics and immunochemical procedures.
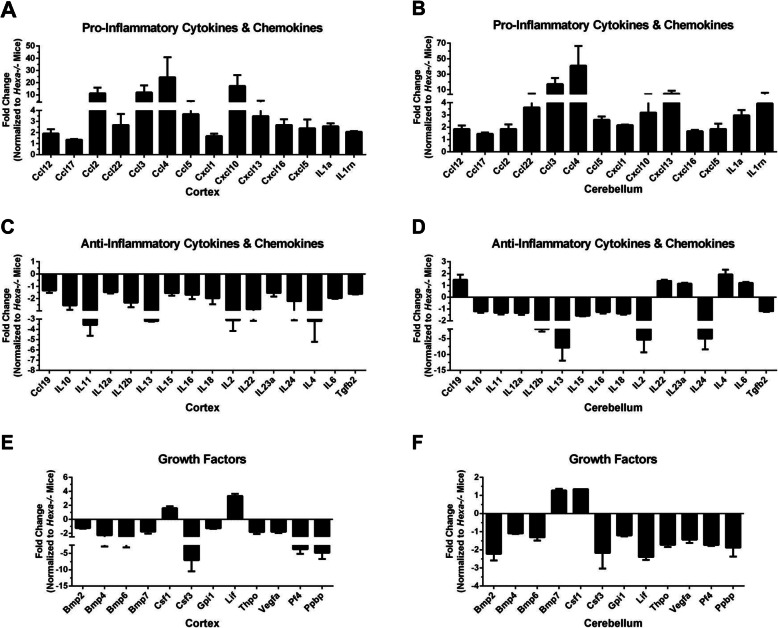
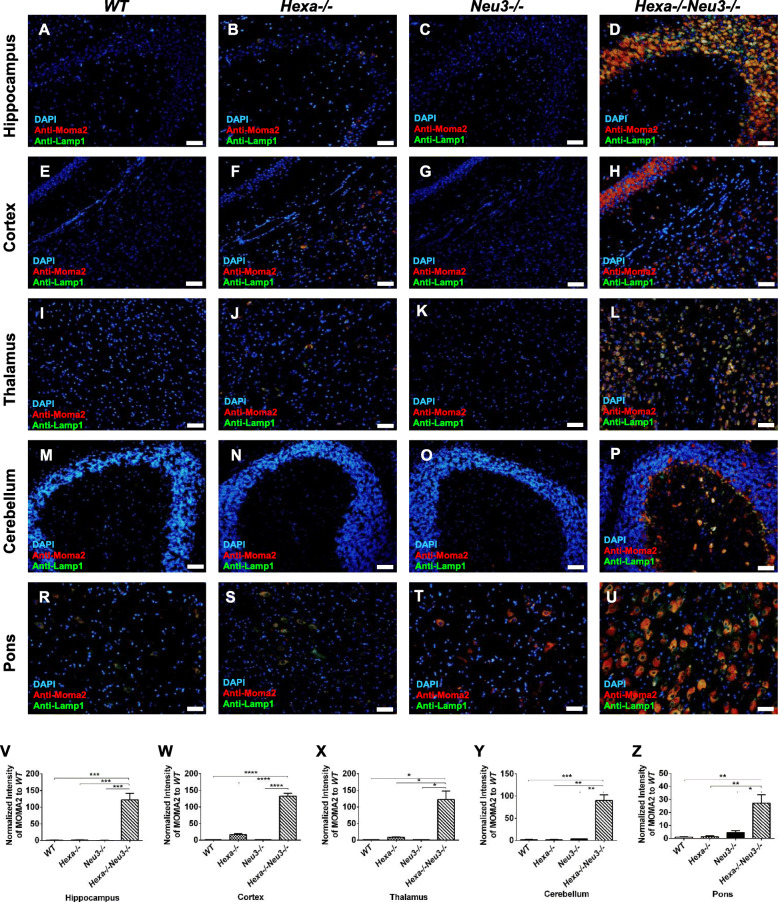
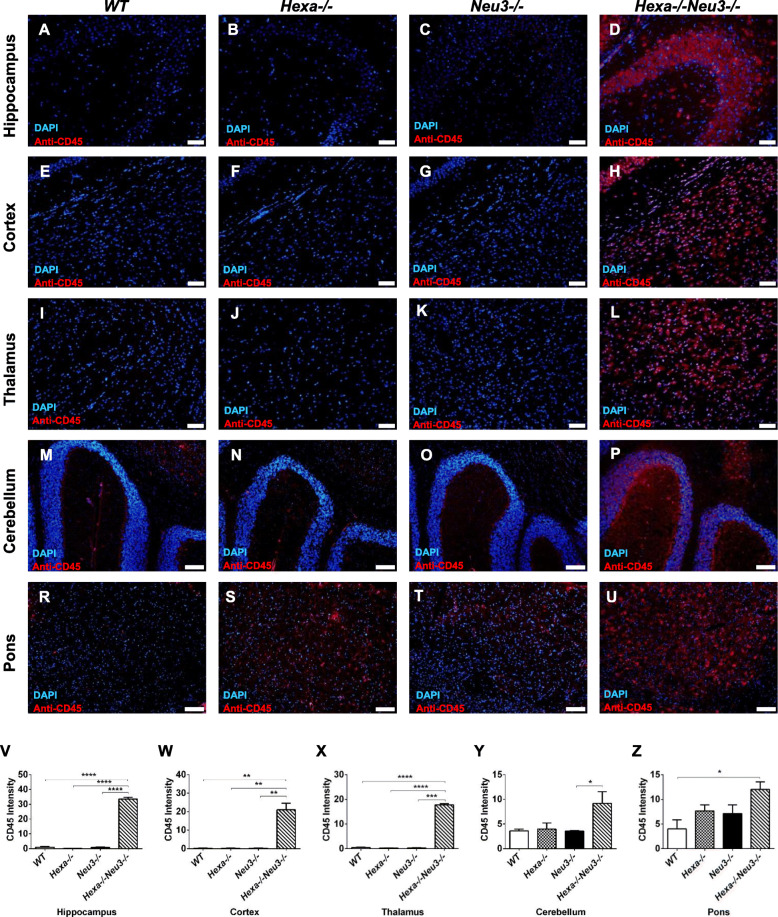
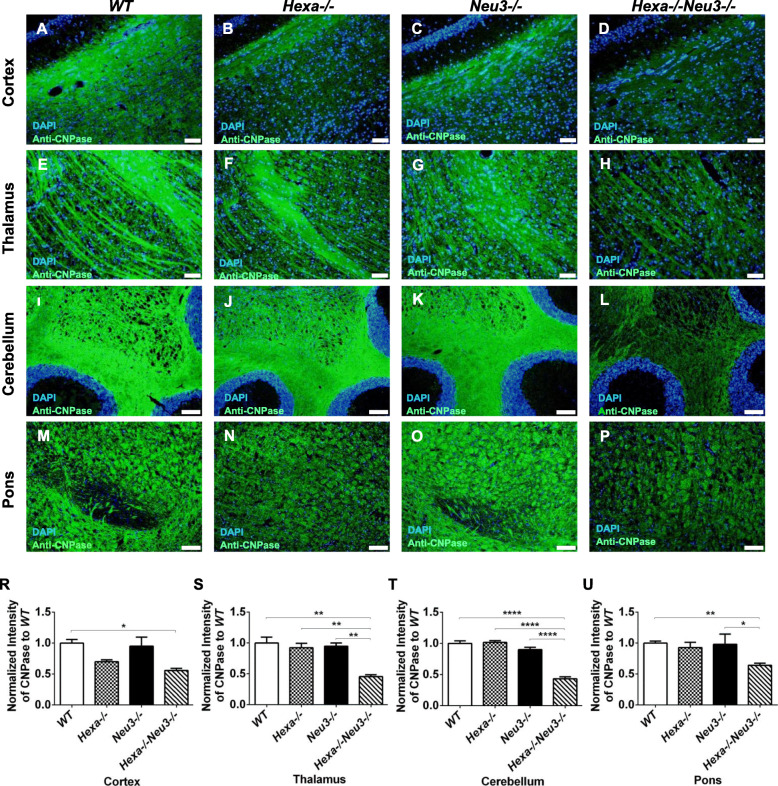
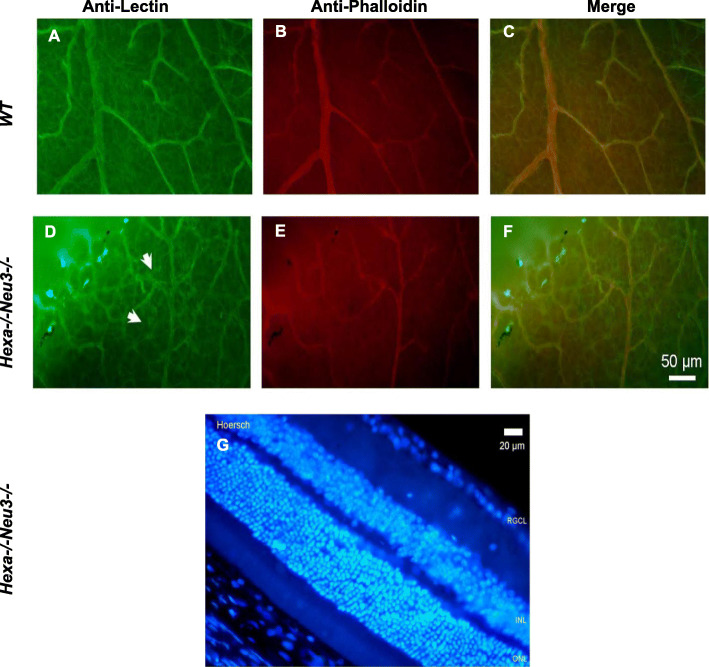
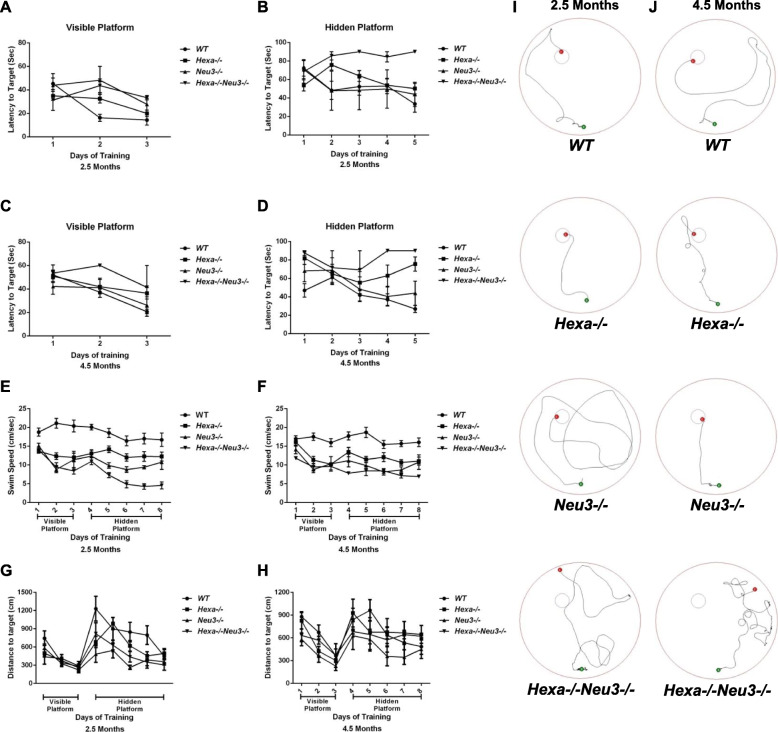
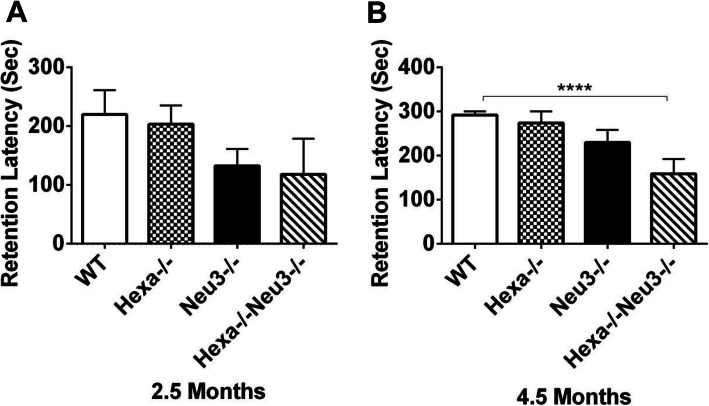
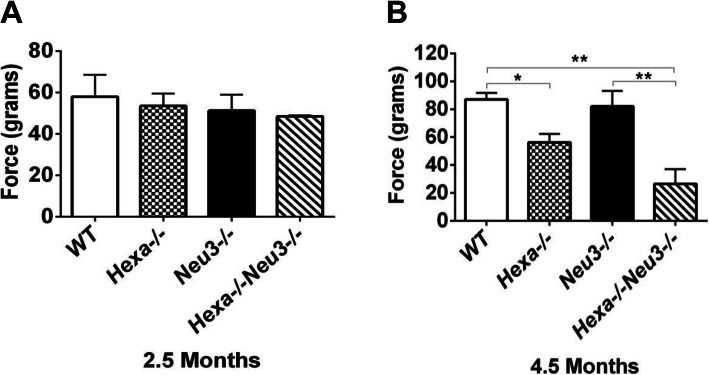
Results: We found elevated levels of pro-inflammatory cytokine and chemokine transcripts, such as Ccl2, Ccl3, Ccl4, and Cxcl10 and also extensive microglial and astrocyte activation and proliferation, accompanied by peripheral blood mononuclear cell infiltration in the vicinity of neurons and oligodendrocytes. Behavioral tests demonstrated a high level of anxiety, and age-dependent loss in both spatial learning and fear memory in Hexa-/-Neu3-/- mice compared with that in the controls.
Conclusion: Altogether, our data suggest that Hexa-/-Neu3-/- mice display a phenotype similar to Tay-Sachs patients suffering from chronic neuroinflammation triggered by GM2 accumulation. Furthermore, our work contributes to better understanding of the neuropathology in a mouse model of early onset Tay-Sachs disease.
Keywords: Behavior; GM2; Mouse model; Neuroinflammation; Tay-Sachs disease.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Yuziuk JA, Bertoni C, Beccari T, Orlacchio A, Wu Y-Y, Li S-C, et al. Specificity of mouse G M2 activator protein and β-N-acetylhexosaminidases A and B. J Biol Chem. 1998;273:66–72. - PubMed
-
- Seyrantepe V, Demir SA, Timur ZK, Von Gerichten J, Marsching C, Erdemli E, et al. Murine sialidase Neu3 facilitates GM2 degradation and bypass in mouse model of Tay-Sachs disease. Exp Neurol. 2017;2018(299):26–41. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
