

Characterization of the genomically encoded fosfomycin resistance enzyme from Mycobacterium abscessus
- PMID: 32952996
- PMCID: PMC7478155
- DOI: 10.1039/c9md00372j
Characterization of the genomically encoded fosfomycin resistance enzyme from Mycobacterium abscessus
Abstract
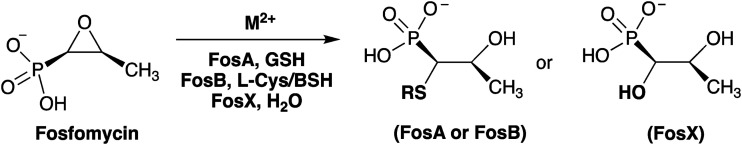
Mycobacterium abscessus belongs to a group of rapidly growing mycobacteria (RGM) and accounts for approximately 65-80% of lung disease caused by RGM. It is highly pathogenic and is considered the prominent Mycobacterium involved in pulmonary infection in patients with cystic fibrosis and chronic pulmonary disease (CPD). FosM is a putative 134 amino acid fosfomycin resistance enzyme from M. abscessus subsp. bolletii that shares approximately 30-55% sequence identity with other vicinal oxygen chelate (VOC) fosfomycin resistance enzymes and represents the first of its type found in any Mycobacterium species. Genes encoding VOC fosfomycin resistance enzymes have been found in both Gram-positive and Gram-negative pathogens. Given that FosA enzymes from Gram-negative bacteria have evolved optimum activity towards glutathione (GSH) and FosB enzymes from Gram-positive bacteria have evolved optimum activity towards bacillithiol (BSH), it was originally suggested that FosM might represent a fourth class of enzyme that has evolved to utilize mycothiol (MSH). However, a sequence similarity network (SSN) analysis identifies FosM as a member of the FosX subfamily, indicating that it may utilize water as a substrate. Here we have synthesized MSH and characterized FosM with respect to divalent metal ion activation and nucleophile selectivity. Our results indicate that FosM is a Mn2+-dependent FosX-type hydrase with no selectivity toward MSH or other thiols as analyzed by NMR and mass spectroscopy.
This journal is © The Royal Society of Chemistry 2019.
Figures







References
-
- Griffith D. E., Aksamit T., Brown-Elliot B. A., Catanzaro A., Daley C., Gordin F., Holland S. M., Horsburgh R., Huitt G., Iademarco M. F., Iseman M., Olivier K., Ruoss S., von Reyn C. F., Wallace, Jr. R. J., Winthrop K. Am. J. Respir. Crit. Care Med. 2007;175:367–416. - PubMed
-
- Adekambi T., Berger P., Raoult D., Drancourt M. Int. J. Syst. Evol. Microbiol. 2006;56:133–143. - PubMed
-
- Griffith D. E., Girard W. M., Wallace, Jr. R. J. Am. Rev. Respir. Dis. 1993;147:1271–1278. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous