

Subcellular Localization And Formation Of Huntingtin Aggregates Correlates With Symptom Onset And Progression In A Huntington'S Disease Model
- PMID: 32954323
- PMCID: PMC7425396
- DOI: 10.1093/braincomms/fcaa066
Subcellular Localization And Formation Of Huntingtin Aggregates Correlates With Symptom Onset And Progression In A Huntington'S Disease Model
Abstract
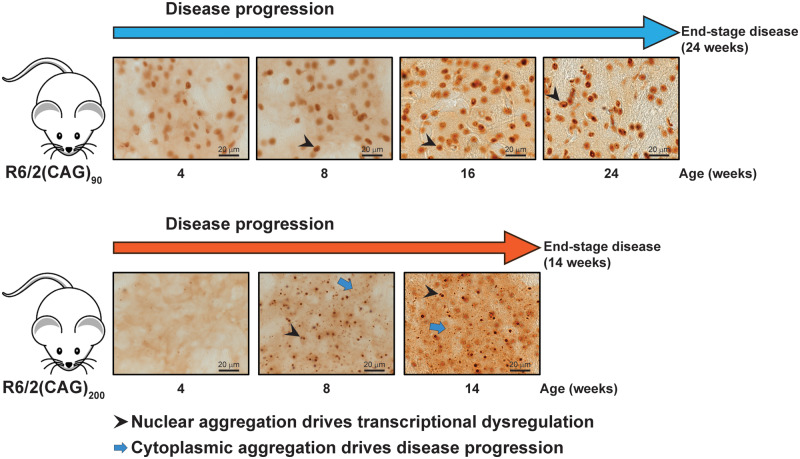
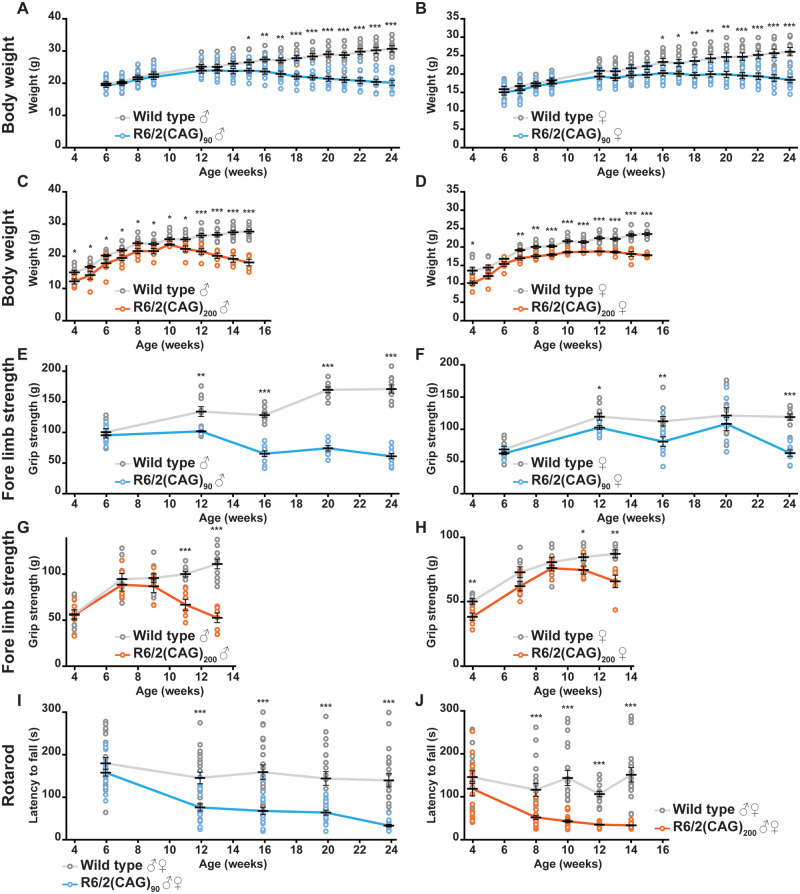
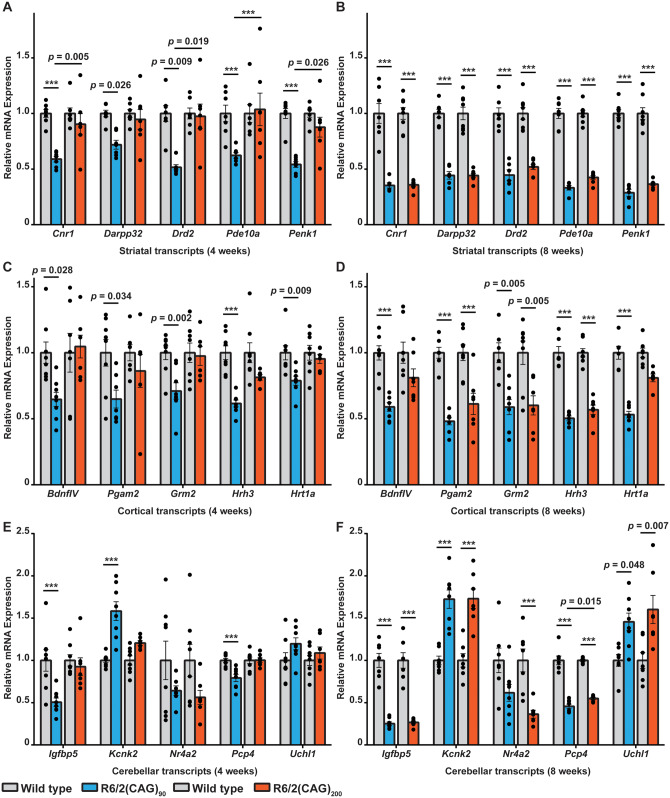
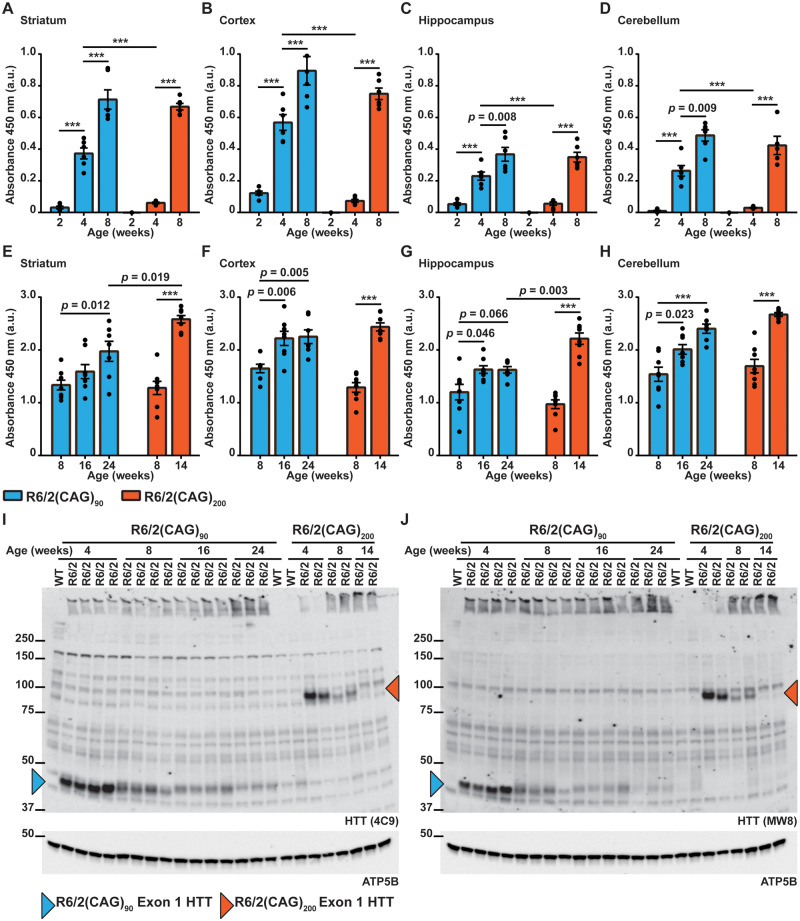
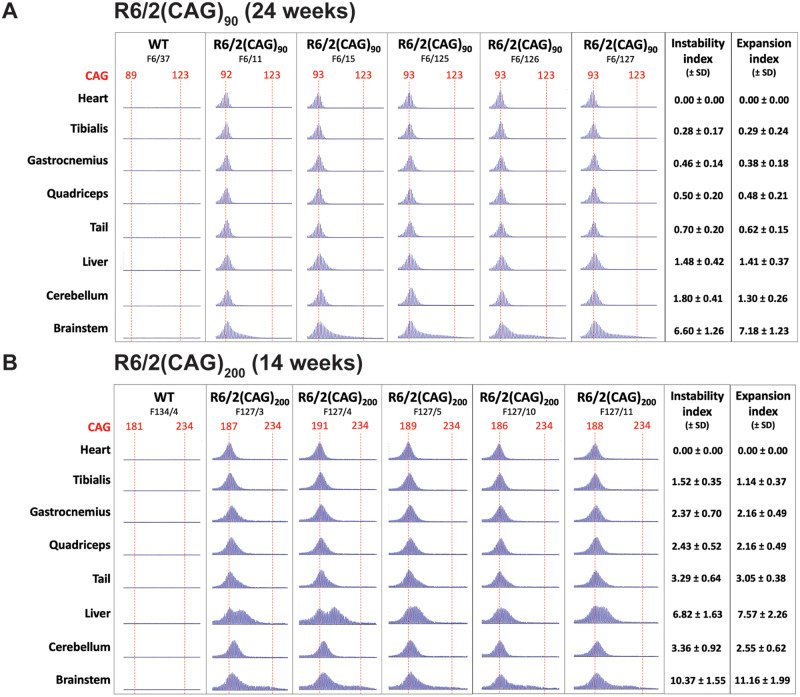
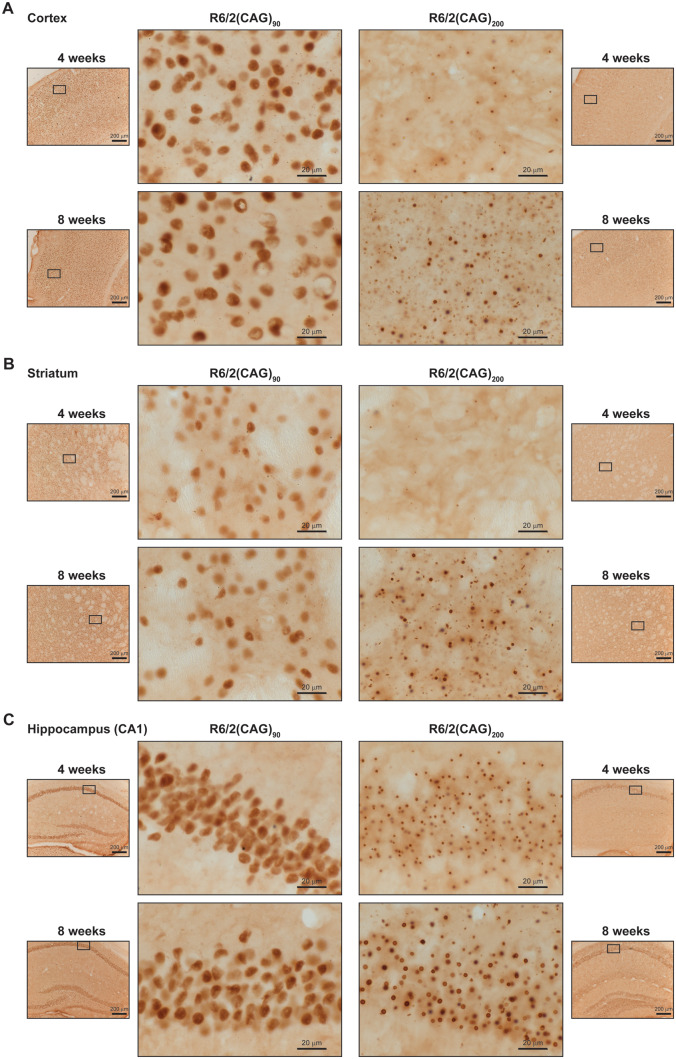
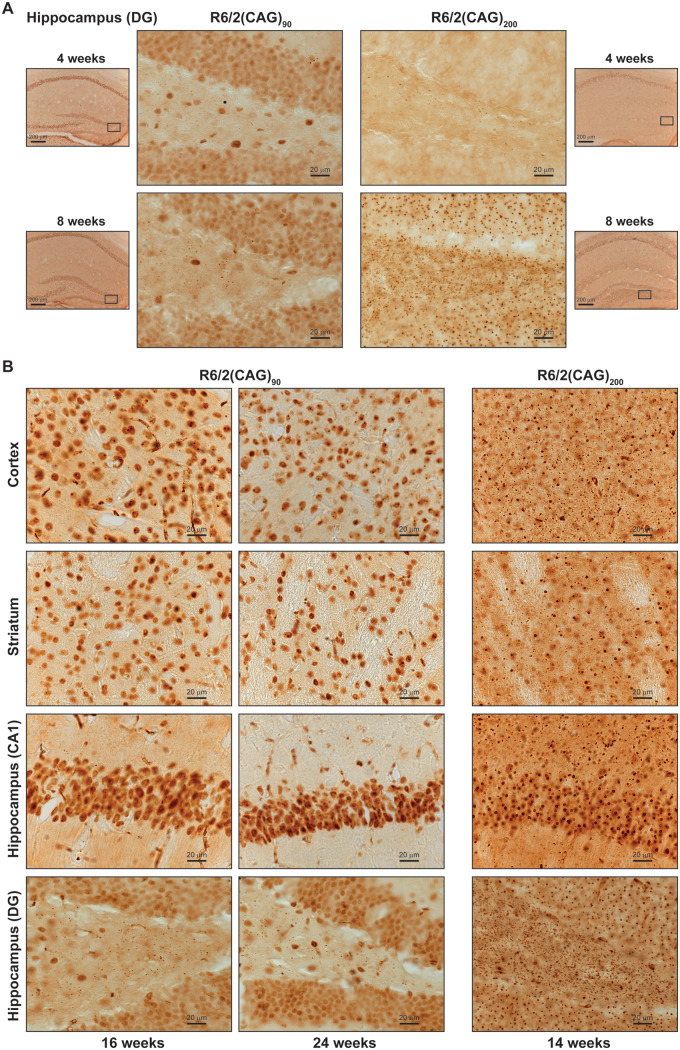
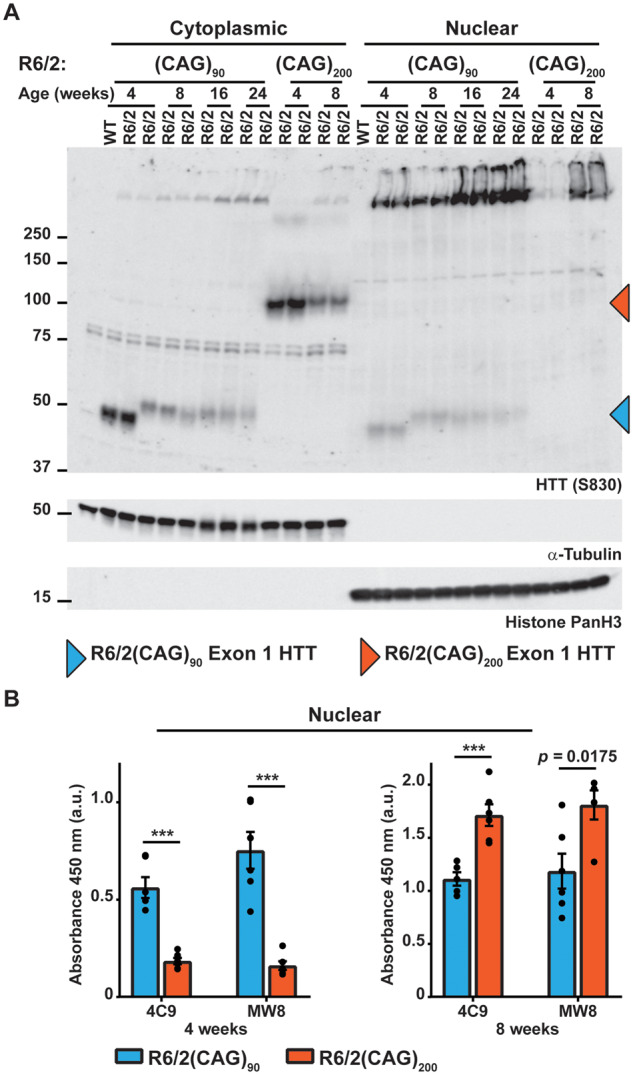
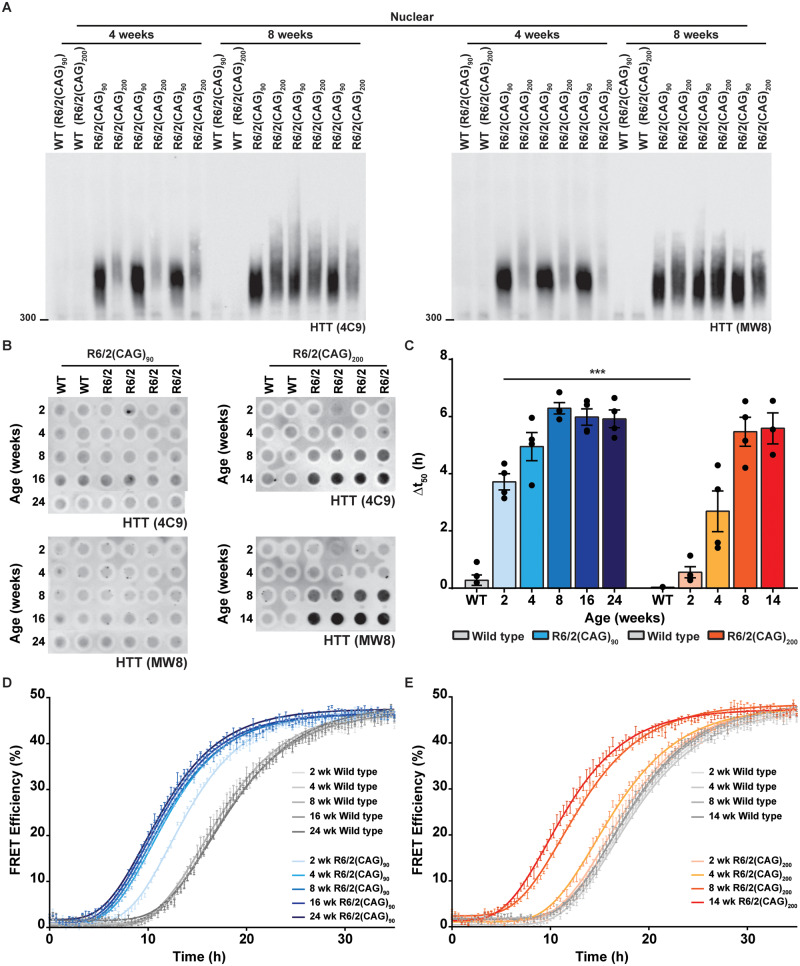
Huntington's disease is caused by the expansion of a CAG repeat within exon 1 of the HTT gene, which is unstable, leading to further expansion, the extent of which is brain region and peripheral tissue specific. The identification of DNA repair genes as genetic modifiers of Huntington's disease, that were known to abrogate somatic instability in Huntington's disease mouse models, demonstrated that somatic CAG expansion is central to disease pathogenesis, and that the CAG repeat threshold for pathogenesis in specific brain cells might not be known. We have previously shown that the HTT gene is incompletely spliced generating a small transcript that encodes the highly pathogenic exon 1 HTT protein. The longer the CAG repeat, the more of this toxic fragment is generated, providing a pathogenic consequence for somatic expansion. Here, we have used the R6/2 mouse model to investigate the molecular and behavioural consequences of expressing exon 1 HTT with 90 CAGs, a mutation that causes juvenile Huntington's disease, compared to R6/2 mice carrying ∼200 CAGs, a repeat expansion of a size rarely found in Huntington's disease patient's blood, but which has been detected in post-mortem brains as a consequence of somatic CAG repeat expansion. We show that nuclear aggregation occurred earlier in R6/2(CAG)90 mice and that this correlated with the onset of transcriptional dysregulation. Whereas in R6/2(CAG)200 mice, cytoplasmic aggregates accumulated rapidly and closely tracked with the progression of behavioural phenotypes and with end-stage disease. We find that aggregate species formed in the R6/2(CAG)90 brains have different properties to those in the R6/2(CAG)200 mice. Within the nucleus, they retain a diffuse punctate appearance throughout the course of the disease, can be partially solubilized by detergents and have a greater seeding potential in young mice. In contrast, aggregates from R6/2(CAG)200 brains polymerize into larger structures that appear as inclusion bodies. These data emphasize that a subcellular analysis, using multiple complementary approaches, must be undertaken in order to draw any conclusions about the relationship between HTT aggregation and the onset and progression of disease phenotypes.
Keywords: Huntington’s disease; huntingtin aggregation and seeding; polyglutamine; somatic CAG instability; transcriptional dysregulation.
© The Author(s) (2020). Published by Oxford University Press on behalf of the Guarantors of Brain.
Figures
References
-
- Ast A, Buntru A, Schindler F, Hasenkopf R, Schulz A, Brusendorf L, et al. mHTT seeding activity: a marker of disease progression and neurotoxicity in models of Huntington’s disease. Mol Cell 2018; 71: 675–88 e6. - PubMed
-
- Bates GP, Dorsey R, Gusella JF, Hayden MR, Kay C, Leavitt BR, et al. Huntington disease. Nat Rev Dis Primers 2015; 1: 15005. - PubMed
-
- Benn CL, Landles C, Li H, Strand AD, Woodman B, Sathasivam K, et al. Contribution of nuclear and extranuclear polyQ to neurological phenotypes in mouse models of Huntington’s disease. Hum Mol Genet 2005; 14: 3065–78. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases